Механизмы трансформации диванов — статьи про мебель на Викидивании
Откроем секрет: нет «плохих» и «хороших» механизмов трансформации, бывает неправильный подход к выбору подходящей модели. Разобраться в том, какие бывают диваны, найти «свой» вариант поможет обзор с короткими иллюстрирующими видео.
Подбирая тип трансформации, обращайте внимание на то, как раскладывается диван, легкость и скорость, соприкосновение с полом и наличие бельевого ящика. Важно и назначение будущего дивана – ежедневное использование или раскладывание от случая к случаю?
Книжка
Механизм, знакомый многим с детства. Трансформация из дивана в кровать проста и интуитивно понятна. Проверенные временем раскладные диваны оснащены вместительным бельевым отсеком под сиденьем и имеют привлекательную цену за счет простоты и неприхотливости конструкции.
Как разложить: сиденье поднимается вверх до щелчка, а затем опускается.![]()
Чем хорош: низкой ценой, быстрым способом раскладывания, вместительным ящиком для белья.
О чем следует помнить: расстояние от спинки до стены должно быть не менее 10 см., для раскладывания необходимо приложить усилия.
Еврокнижка
Надежный и простой в использовании механизм трансформации дивана. Благодаря отсутствию петель на стыке спальная поверхность получается ровной и удобной. Спальное место располагается вдоль дивана, что идеально для узких помещений. Не нужно задумываться, как разложить или сложить «еврокнижку» и нет необходимости отодвигать каждый раз от стены – конструкция модели выдвигается вперед.
Как разложить: Выкатите сиденье и опустите спинку – место для отдыха готово.
Чем хорош:
надежностью и вместительным ящиком для белья, нет необходимости двигать диван каждый раз при раскладывании, подходит для узких комнат.О чем следует помнить: модели без колес могут царапать пол.
Пантограф
Данная разновидность имеет все преимущества «еврокнижки», и даже больше. Поднимающиеся диваны не оснащены роликами и раздвигаются не касаясь пола. Подъемный механизм выталкивает сиденье вверх и оно словно шагает вперед, оставляя напольное покрытие нетронутым. Обратный механизм складывания также не требует усилий.
Как разложить: потяните вверх и на себя сиденье и опустите спинку.
Чем хорош: не взаимодействует с полом, оснащен бельевым отсеком, не нужно двигать при раскладывании.
О чем следует помнить:
Еврософа
Системы раскладывания на металлическом каркасе отличаются наждежностью и прочностью. Отлично подходят для активной ежедневной эксплуатации, с легкостью перенесут перестановки и переезды.
Как разложить: надавите сверху вниз на край спинки – плавно опускаясь, она займет горизонтальное положение. Для обратной трансформации потяните раму спинки вверх.
Чем хорош: имеет в основании металлический каркас, не требует отодвигания дивана от стены.
О чем следует помнить: единственный недостаток таких систем — относительно высокая стоимость.
Дельфин
Диван, в составе которого находится механизм «дельфин», раскладывается с особой легкостью и скоростью. Устройство получило название в честь одноименного животного благодаря схожести движений.
Как правило, встречается в угловых диванах. Каркас спального места удерживается за счет специальных кронштейнов на уровне основной части дивана. Благодаря компактности такая модель удачно впишется в интерьер небольшой комнаты, а вместительный ящик для белья под угловой частью поможет спрятать домашний текстиль и бытовую мелочь.![]()
Как разложить: выдвините из-под сиденья раму и потяните вверх.
Пума
Подобно кошачьей поступи, диваны с механизмом «пума» раскладываются максимально тихо и аккуратно. Никаких царапин от ножек или промятых дорожек от колесиков – даже самое требовательное напольное покрытие не пострадает при ежедневном раскладывания. При трансформации нужны минимальные усилия: немного потяните сиденье вверх, а дальше всю работу сделает механизм.
Как разложить: потяните спальное место на себя за сиденье, а следом за ним появится часть, расположенная внизу.
Чем хорош: бесшумностью и бережным отношением к напольному покрытию, легко раскладывается.
О чем следует помнить: у прямых моделей нет ящика для белья. У угловых моделей он находится за оттоманкой.
Аккордеон
Механизм «аккордеон» получил свое название благодаря сходству раскладывания с движением мехов музыкального инструмента. Спальное место получается ровным и широким, без ощутимых стыков. Модель относится к категории раздвижных диванов и ощутимо съедает пространство в длину, выбирайте ее если пространство позволяет подобный простор для манипуляций.
Спальное место получается ровным и широким, без ощутимых стыков. Модель относится к категории раздвижных диванов и ощутимо съедает пространство в длину, выбирайте ее если пространство позволяет подобный простор для манипуляций.
Как разложить: потяните петлю и сиденье поедет вперед, а спинка развернется и разложится на две части.
Чем хорош: легкостью раскладывания.
О чем следует помнить: требует много места для раскладывания, поэтому не ставьте перед сиденьем столик или другую мебель. За механизмом нужно ухаживать и регулярно смазывать.
Караван
Механизм раскладывания на металлокаркасе, надежный и долговечный. За счет трансформации вперед организует широкое спальное место (аналог двуспальной кровати) при сравнительно небольших размерах дивана «караван».
Как разложить: потяните сиденье вверх – оно начнет описывать в воздухе дугу. В это же время из-под основания друг за другом появятся две дополнительные платформы. Когда сиденье опуститься на опоры, три функциональные части образуют ровную поверхность для сна.
В это же время из-под основания друг за другом появятся две дополнительные платформы. Когда сиденье опуститься на опоры, три функциональные части образуют ровную поверхность для сна.
О чем следует помнить: так как место под сиденьем «каравана» занято дополнительными платформами, у диванов данного типа нет бельевого ящика.
Французская раскладушка
Французская раскладушка или «седафлекс» – легкое и компактное европейское гостевое решение. Из-за особенностей конструкции модель раскладывается значительно дольше «пумы» или «еврокнижки», в ней нет бельевого ящика, а также такой диван не подходит для ежедневного раскладывания. Если вам требуется вариант для частого использования, ищите модель французской раскладушки, где в поддерживающем основании вместо ткани деревянный каркас – этот вариант более полезен для позвоночника.
Как разложить: поднимите верхнюю подушку-сиденье и потяните за ножки вперед – спальное место, сложенное гармошкой, скрывается под сиденьем.
Чем хорош: легкостью и прочностью конструкции.
О чем следует помнить: нет ящика для белья, слишком мягкая постель (не рекомендуется для ежедневного сна), диваны с раскладывающимся вперед механизмом не всегда удачно вписываются в узкие помещения.
Рич
Механизм «Рич» или «Спартак» – американская раскладушка двойного сложения. В его конструкции – металлическая рама и сварная решетка, надежная и долговечная. Система подходит как для гостевого, так и для ежедневного использования. Главная особенность механизма – возможность «убирать» сиденье под спальное место, что автоматически облегчает задачу трансформации.
Как разложить: поднимите сиденье и потяните систему на себя – спальное место, сложенное вдвое, появится из-под основания.
Чем хорош: легко раскладывается, надежен и долговечен.
Все категории и типы диванов имеют существенные и принципиальные различия. Отнеситесь к выбору модели с особой внимательностью и наслаждайтесь комфортом каждый день.
Механизмы диванов — Обзор видов и типов механизмов трансформации диванов
Диваны стали действительно незаменимой мебелью в наших домах. Они не только обеспечивают уютное место для отдыха в кругу семьи, но и представляют собой в разложенном виде полноценное спальное место. Чтобы трансформировать диван было как можно проще, конструкторы мебели разработали специальные механизмы. Они отличаются как по способу раскладывания, так и по другим характеристикам: удобству для ежедневного сна, занимаемой площади в собранном и разобранном виде и прочим.
Какие механизмы диванов бывают? Чем они отличаются между собой? Какой механизм дивана лучше для сна? Разберемся вместе.
Механизмы диванов по типу трансформации
Существуют следующие механизмы раскладывания:
- американская раскладушка;
- аккордеон;
- бриз;
- еврокнижка;
- клик-кляк;
- еврософа;
- танго;
- сезам;
- юниор;
Рассмотрим подробнее данные типы раскладывания диванов.![]()
Американская раскладушка
Очень простой в обращении механизм, разложить такой диван сможет даже школьник. Необходимо потянуть спинку на себя ー спальное место своеобразным образом свернуто внутри. При небольшом усилии оно развернется ー сначала ножками вверх, его необходимо раскатать дальше ー и соберется в полноценную кровать.
Еще одно ценное преимущество такого механизма, помимо легкого раскладывания, заключается в удобстве спального места. Оно не имеет швов или стыков элементов дивана, характерных для других видов трансформации. Вы спите на полноценном матрасе.
Этот вид трансформации подойдет как для ежедневного сна одного или двух человек, так и для использования такого дивана в качестве гостевого. Мебель «американская раскладушка» очень компактна ー органично впишется даже в небольшое пространство гостиной.
Аккордеон
Ставший уже классическим для наших квартир механизм «аккордеон» по праву заслуживает своей славы.![]() Это действительно простой и удобный в эксплуатации тип раскладывания.
Это действительно простой и удобный в эксплуатации тип раскладывания.
Диван складывается в виде гармошки, состоящей из 3 частей: сиденья, передней и задней части спинки. В разложенном виде они становятся соответственно изножьем, средней частью спального места и изголовьем.
Особенностью этого типа является то, что при раскладывании образуется действительно просторное спальное место. На таком диване могут спокойно спать 2 взрослых и 1 ребенок! Стыки в элементах подогнаны достаточно плотно ー они практически не ощущаются.
В сложенном виде диван-аккордеон компактен, а вот в разложенном потребует много места. Обеспечьте примерно 2 на 1,5-2 метра площади перед ним.
Бриз
Удобный в использовании механизм, суть которого состоит в выкатывании части спального места вперед. Трансформация достаточно простая ー легко производится одной рукой. Не требует отодвигания дивана от стены и не занимает много места при раскладывании.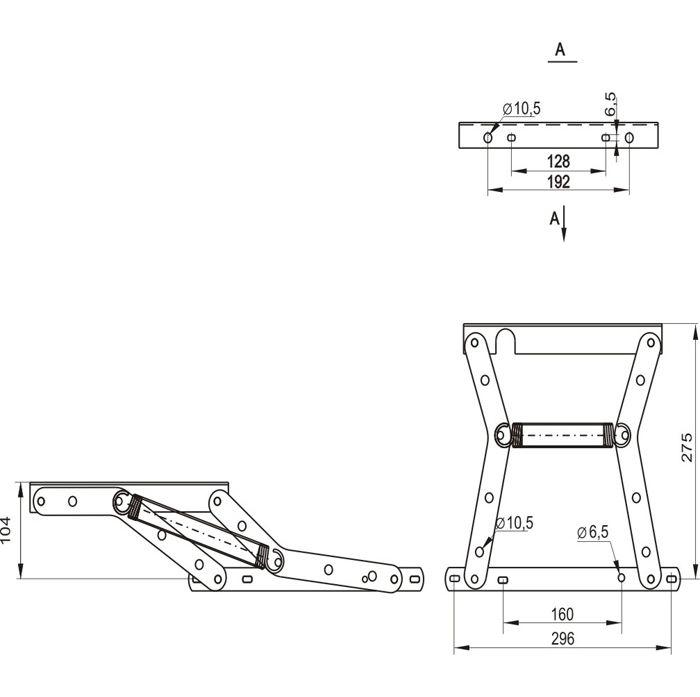
Спальное место получается вместительным, могут спать 2 взрослых человека. Однако для ежедневного сна может быть не очень удобным, так как механизм бриз предполагает несколько продольных и поперечных стыков.
Диван с подобным типом трансформации отлично подойдет для гостиной в качестве мебели для отдыха и дополнительного спального места.
Еврокнижка
Диван с этим механизмом не нужно отодвигать от стены, чтобы разложить. Его трансформация проводится в 2 действия ー сначала передняя часть выкатывается вперед, а затем в образовавшееся пространство укладывается спинка.
Спальное место имеет только 1 стык, что делает его крайне удобным для ежедневного сна. На диване в разложенном виде могут спать 2 взрослых человека ー он достаточно просторный. Другие преимущества этой модели:
- на сложенном диване 1 человек может лечь в полный рост;
- занимает мало места при раскладывании.


У диванов с подобным механизмом достаточно большая длина в сложенном состоянии, из-за чего они могут быть неуместны в малогабаритных квартирах. Зато даже в сложенном виде на таком диване уместится вся семья для отдыха перед телевизором.
Клик-кляк
Этот механизм предполагает раскладывание дивана по типу книжки. Производится в 2 действия ー сначала сиденья надо слегка приподнять, а затем опустить ー спинка также примет горизонтальное положение. Обратите внимание, что в процессе трансформации потребуется немного места у стены.
В разложенном виде представляет собой полноценное спальное место для 2 человек. Нет стыков или перепадов высоты, что делает клик-кляк одним из самых удобных механизмов для полноценного сна на каждый день.
Стоит также отметить, что диваны с таким механизмом оборудованы вместительными ящиками для белья.
Еврософа
Интересный по своему исполнению механизм трансформации, заключающийся в 2 действиях:
- выдвигается ящик для белья;
- на него как на опору раскладывается половинка сиденья.

Модели с таким механизмом раскладывания компактны в сложенном виде, занимают немного полезной площади комнаты. Спать на них очень удобно ー стык двух половинок практически не ощущается, а материал сидений упругий и в меру жесткий.
Танго
Этот механизм схож с системой трансформации клик-кляк. Для того чтобы из дивана сделать спальное место, сиденье надо приподнять вверх до определенной точки, а затем опустить. Спинка опустится одновременно с сиденьем, образуя ровное пространство для сна или отдыха. Отличие механизма танго ー в растровых подлокотниках, которые можно поднять под нужным вам углом. Во время сна можно сделать удобный подголовник.
Такие диваны подходят для сна вдвоем, их часто покупают в детскую комнату. Обратите внимание ー для них понадобится чуть больше места, чем, к примеру, для мебели с системой трансформации аккордеон.
Сезам
Этот механизм ー авторская разработка специалистов фабрики ANDERSSEN и компании Steelglass.![]() Он создан в соответствии с запросами клиентов на удобный и многофункциональный диван для гостиной или детской. Суть механизма в том, что он раскладывается в односпальную кровать с удобным матрасом, уложенным на ортопедические латы. Трансформация проводится в 2 действия:
Он создан в соответствии с запросами клиентов на удобный и многофункциональный диван для гостиной или детской. Суть механизма в том, что он раскладывается в односпальную кровать с удобным матрасом, уложенным на ортопедические латы. Трансформация проводится в 2 действия:
- выдвигается за низ спрятанная в глубине кушетки секция;
- поднимается спальное место с высоким изголовьем.
Эту систему уже оценили наши покупатели ー диваны с механизмом сезам активно заказывают для детской или комнаты подростка. Вариант также удобен в качестве дополнительного места в случае ночевки гостя.
Юниор
Это идеальное решение для детской комнаты малой площади. В собранном виде мебель с таким механизмом представляет собой обычный диван. В разобранном ー 2 односпальные кровати, отделенные друг от друга мягкой секцией. Трансформация осуществляется очень легко ー достаточно вытянуть за петлю сиденье, а затем опустить спинку на место изголовья.
Кровати подходят для ежедневного сна, так как не имеют перепадов высоты и характеризуются умеренной жесткостью. Это замечательный выбор для спальни подросших детей или для гостиной в качестве дополнительных спальных мест.
Виды механизмов в зависимости от конструкции диванов
На сегодняшний день на рынке мягкой мебели представлены диваны различных конструкций с характерными для них типами раскладывания. Рассмотрим наиболее популярные из них:
- прямые модели ー обычно они имеют механизмы трансформации еврокнижка, клик-кляк, аккордеон, американская раскладушка, танго и другие. Такие диваны компактны в сложенном виде и достаточно вместительны в разложенном;
- угловые диваны обычно оснащаются такими механизмами трансформации как аккордеон, бриз, американская раскладушка, нью-роллер и еврософа. Они удобны для отдыха всей семьей перед телевизором, но занимают больше площади комнаты, чем прямые модели;
- кресла-кровати образуют полноценные спальные места для 1 человека благодаря механизмам трансформации аккордеон, нью-роллер или юниор. Отличное решение проблемы дополнительного спального места в небольших помещениях;
- кушетки представляют собой диваны, которые можно трансформировать в спальное место для 1 человека. Идеально подходят для детских комнат или малогабаритных квартир.
Лучшие механизмы диванов для сна и отдыха
Если диван будет вами использоваться и в качестве места для ежедневного сна, подойти к его выбору надо ответственно. Ведь он должен отвечать не только эстетическим, но и некоторым другим требованиям, например, иметь ортопедическое основание, при раскладывании образовывать минимум стыков и прочее.
Какой механизм дивана лучше для ежедневного сна? Мы подготовили топ-6 видов трансформации, которые идеально подойдут для полноценного отдыха:
- Американская раскладушка. Идеален для сна за счет спального места без стыков и удобного основания кровати.
- Еврософа. Удобный механизм трансформации, обеспечивающий вместительное спальное место. Получается поверхность только с 1 стыком, который не ощущается в процессе сна. Хороший вариант для основного варианта спального места.
- Клик-кляк. Простой в раскладывании механизм. Стыков в разложенном виде у таких диванов нет, что крайне удобно для сна каждую ночь. Единственный нюанс ー кого-то может смутить отсутствие бортика в изголовье.
Выбор механизма трансформации дивана в зависимости от потребности
Если вы ищете диван, который в разобранном виде был бы очень просторным, советуем обратить внимание на следующие виды трансформации:
Если вам нужен как можно более компактный вариант мебели, купите его с одним из таких видов раскладывания как:
Наиболее удобные диваны для детской ー это:
- юниор;
- сезам;
- кушетка.
Какой бы диван фирмы ANDERSSEN вы ни выбрали, будьте уверены ー он прослужит вам много лет, оставаясь таким же удобным и привлекательным с виду. Мебель нашей фабрики создает в доме уют и тепло.
Мебель нашей фабрики создает в доме уют и тепло.
Механизм трансформации диванов Аккордеон | Фабрика мягкой мебели ANDERSSEN
Механизм-аккордеон
Вы ищете максимально удобный механизм, который можно было бы разбирать и собирать ежедневно? Если да, то вас должен обязательно заинтересовать механизм «Аккордеон». Такие модели предлагает мебельная фабрика «Андерссен», основное направление деятельности которой – производство и продажа мягкой мебели.
«Аккордеон» – механизм на металлокаркасе, основной частью спального места которого становится спинка. Данный механизм, благодаря своим конструкционным особенностям, отличается высокой надежностью и продолжительным сроком службы. Он стоит из трех каркасов, упругость которых обеспечивается деревянными ламелями. Диваны с механизмом аккордеон очень просто раскладывать, превращая диван в кровать с комфортным ровным спальным местом.
Процесс трансформации заключается в перемещении сиденья вперед. Вслед за сиденьем двигается, опускаясь и раскладываясь, спинка. Превращение кровати в диван осуществляется в обратном порядке. В разобранном состоянии такая мебель обеспечивает очень ровное спальное место, которое помогает достичь максимального расслабления позвоночника и мышц спины во время отдыха.
Вслед за сиденьем двигается, опускаясь и раскладываясь, спинка. Превращение кровати в диван осуществляется в обратном порядке. В разобранном состоянии такая мебель обеспечивает очень ровное спальное место, которое помогает достичь максимального расслабления позвоночника и мышц спины во время отдыха.
Мягкая мебель с механизмом «Аккордеон», благодаря использованию в конструкции специальных деревянных ламелей, обладает хорошим ортопедическим эффектом. Многие предпочитают купить механизм-аккордеон в связи с тем, что он позволяет эффективно экономить полезное пространство комнаты.
Купить механизм аккордеон мебельной фабрики Anderssen
Если вас интересуют механизмы «Аккордеон» от производителя, обратите внимание на продукцию мебельной фабрики «Андерссен». Широчайший ассортимент моделей позволит подобрать вариант, который будет соответствовать вашим требованиям к габаритам (представлены как большие, так и малые механизмы), типу и цвету обивки, функциональности и цене.![]() Ухаживать за мебелью «Андерссен» не составляет труда. Производитель выпускает механизмы со съемными чехлами, которые очень просто чистить и стирать.
Ухаживать за мебелью «Андерссен» не составляет труда. Производитель выпускает механизмы со съемными чехлами, которые очень просто чистить и стирать.
Для тех, кто желает подобрать механизм-аккордеон, фото моделей представлены в каталоге. Качественные изображения помогут вам определиться с выбором варианта, который идеально впишется в интерьер вашей квартиры. В каталоге указаны технические характеристики, вариант обивки, в котором выполнен механизм-аккордеон, цена изделия.
Фабрика ANDERSSEN производит широкий модельный ряд угловых диванов с механизмом аккордеон.
Если у вас возникли вопросы или вы не нашли в каталоге понравившуюся модель, свяжитесь с нашими консультантами по телефонам: 8 (495) 601-96-45 (Москва и область), +7 (499) 112-45-95 (регионы). Специалисты ответят на все ваши вопросы.
Механизм трансформации диванов Еврокнижка | Фабрика мягкой мебели ANDERSSEN
Механизм еврокнижка
Механизм трансформации «Еврокнижка» фабрика изготавливает самостоятельно исключительно из экологически чистых материалов таких, как сосновый брус, фанера, ламинированное ДСП.![]() Дополнительную устойчивость модели в целом обеспечивает используемый в каркасе массив бука.
Дополнительную устойчивость модели в целом обеспечивает используемый в каркасе массив бука.
На диване с механизмом евроскнижка очень удобно отдыхать, сидя в компании с любимым фильмом и мягким пледом. Удобство посадки обеспечивается эргономичными подлокотниками, которые повторяют изгиб руки, и большими мягкиеми подушками-спинками. Отдохнуть после тяжелого трудового дня можно, просто присев на диван и откинув голову назад.
Диван с механизмом «Еврокнижка» имеет небольшую глубину, но на нем можно удобно прилечь, не раскладывая. Для того, чтобы на диване смог комфортно разместиться один человек, достаточно просто убрать подушки-спинки.
Для того, чтобы диван-еврокнижка превратился в полноценную двуспальную кровать с широким ровным спальным местом, необходимо выполнить 2 простых действия: переместить вперед выкатную часть и опустить спинку дивана. Легкость скольжения выкатной части дивана обеспечивают двойные ролики из обрезиненного пластика и тщательно обработанные направляющие из бука.
Комфорт спального места достигается, благодаря высокоэластичному ППУ плотностью 30кг/м³ и высотой 12см. При трансформации дивана со стороны стены остается соединительная спинка, которая сохраняет внешний вид обоев и придает эстетичный внешний вид спальному месту.
Бельевой ящик в диванах-еврокнижках ANDERSSEN поистине считается предметом гордости конструкторов фабрики. Он больше ящиков обычных еврокнижек и не разделен ригелем. Сам ящик выполнен из ламинированной ДСП, по торцевому краю обклеен кромкой ПВХ, что позволяет легко поддерживать чистоту. Дно выполнено также из ламинированной ДСП, что повышает его прочность – ящик выдерживает нагрузку до 200 кг!
Если у вас возникли вопросы или вы не нашли в каталоге понравившуюся модель, свяжитесь с нашими консультантами по телефонам: 8 (495) 601-96-45 (Москва и область), +7 (499) 112-45-95 (регионы) . Специалисты ответят на все ваши вопросы.
Механизм трансформации диванов Еврософа | Фабрика мягкой мебели ANDERSSEN
Механизм еврософа
«Еврософа» от фабрикиANDERSSEN – это комфортабельный и надежный диван-кровать на деревянном каркасе – удобный и долговечный. Это авторская разработка конструкторов и дизайнеров нашей фабрики.
Это авторская разработка конструкторов и дизайнеров нашей фабрики.
Диван-еврософа – прекрасный вариант для людей, ценящих практичную и функциональную мебель. Модели с механизмом раскладывания «Еврософа» обладают целым рядом преимуществ:
1. Удобство. Днем такое изделие занимает небольшую площадь и при этом позволяет отдыхать с комфортом. Глубина дивана в собранном виде составляет всего 0,9 м. А ночью диван-еврософа оказывается удобной кроватью с ровным спальным местом без стыковочных швов достаточного размера для двоих человек.
2. Надежность. Диваны с таким механизмом трансформации отличаются удивительной прочностью. Только крышка бельевого ящика выдерживает вес до 250 кг!
3. Стиль. Фабрика «Андерссен» предлагает огромный ассортимент моделей и вариантов декора диванов, оснащенных механизмом «Еврософа».
Механизм «еврософа» имеет два уровня раскладывания – положение «диван» и положение «кровать». Конструктивной особенностью дивана является выкатной вместительный бельевой ящик, на который перекидывается половина спального места.![]() Бельевой ящик имеет две открывающиеся крышки, фиксируемые при помощи газового амортизатора, что делает его использование еще более удобным.
Бельевой ящик имеет две открывающиеся крышки, фиксируемые при помощи газового амортизатора, что делает его использование еще более удобным.
Настил спального места дивана выполняется только из высокоплотного пенополиуретана (ППУ) марок S 30/35 и S 30/40. Матрас состоит из двух марок ППУ (толщиной 120 мм и 100 мм) и обеспечивает комфортный отдых. В разложенном виде матрас образует ровное спальное место без стыковочных швов.
Купить механизм еврософа
Диван-еврософа — красивая, качественная и доступная мебель. Купить такое изделие можно винтернет-магазине фабрики мебели Anderssen.
Недавно фабрика мягкой мебели ANDERSSEN начала производить и раскладыне угловые диваны с этим популярным механизмом.
Если у вас возникли вопросы или вы не нашли в каталоге понравившуюся модель, свяжитесь с нашими консультантами по телефонам: 8 (495) 601-96-45 (Москва и область), +7 (499) 112-45-95 (регионы)
. Специалисты ответят на все ваши вопросы.
Механизмы трансформации диванов и кресел
В статье представлены основные особенности различных механизмов трансформации:
«СЕДАФЛЕКС»
Один из самых популярных механизмов трансформации из-за его простоты и надежности. Подходит для ежедневного использования, при разложении образует удобное, ровное спальное место повышенной комфортности. По праву называется «бельгийской кроватью», механизм разработан, запатентован и выпускается франко-бельгийской фирмой SEDAC-MERAL. Максимальная нагрузка на механизм 240кг. Бельевого ящика нет. Раскладка и сон по направлению от стены. Данный механизм трансформации используется в следующих диванах: «Рафаэль».
ПРЕИМУЩЕСТВА:
- Механизм компактный и прячется в диване, позволяя совместить интересный дизайн, комфортную посадку и наличие полноценного спального места.
- Отличительная особенность: наличие пружинного матраса толщиной 10-12 см. Ровное спальное место без стыков.
- В основании спального места под матрасом ортопедические березовые латы, либо металлическая сетка из стали большого сечения, именно это позволяет сравнивать механизм с кроватью. В зоне ног – настил из резинотканевых ремней, который в сложенном виде становится основанием для подушек сиденья и залогом истинного комфорта посадки.
- Оптимальная высота спального места (45 см от пола).
- Центральной опорой спального места служит усиленная стальная «нога», которая и обеспечивает максимальную надежность в эксплуатации.
- Механизм предназначен для ежедневного использования.
КАК РАСКЛАДЫВАТЬ:
Снять подушки сиденья, потянуть механизм вверх и на себя, развернуть звенья, при трансформации становится на стальные ножки – диван разложен.
«МИКСОТОЙЛ»
«Миксотойл» («Французская раскладушка») – механизм, используемый как «гостевой вариант» спального места. Бельевого ящика нет. Максимальная нагрузка на механизм 180 кг. Раскладка и сон по направлению от стены.
Раскладка и сон по направлению от стены.
ПРЕИМУЩЕСТВА:
- Механизм компактный и прячется в диване, позволяя даже в небольшом диване встроить дополнительное гостевое спальное место.
- Поролоновый беспружинный матрац толщиной 5-6 см имеет съемный чехол, что существенно облегчает уход за изделием. Со временем, возможна замена матраса и чехла.
- Гигиеничность спального места (сидим на обивке дивана, а спим на матрасе).
- В основании спального места ортопедические березовые ламели и натяжной элемент в виде полипропиленового тента.
КАК РАСКЛАДЫВАТЬ:
Снять подушки сиденья, потянуть механизм вверх и на себя, развернуть звенья, при трансформации становится на стальные опоры – диван разложен.
«ИФАГРИД» («СПАРТАК»)
Механизм похожий на «Миксотойл» («французская раскла душка»), но спроектированный для ежедневного использования. Производится франко-бельгийской компанией «Седак Мераль» и по лицензии несколькими дочерними предприятиями на территории России. Максимальная нагрузка на механизм 240 кг. Бельевого ящика нет. Раскладка и сон по направлению от стены. Данный механизм трансформации используется в следующих диванах: Lofft, Maxbon, «Брайтон», «Велидж», «Жаклин», «Фьюжн», «Флерон», «Шерон».
Максимальная нагрузка на механизм 240 кг. Бельевого ящика нет. Раскладка и сон по направлению от стены. Данный механизм трансформации используется в следующих диванах: Lofft, Maxbon, «Брайтон», «Велидж», «Жаклин», «Фьюжн», «Флерон», «Шерон».
ПРЕИМУЩЕСТВА:
- Усиленный стальной каркас. Основание спального места — сетчатая стальная решетка.
- Не имеет деревянных и пластиковых соединений, за счет этого обеспечивается надежность и долговечность.
- Поролоновый беспружинный матрас толщиной 7 см имеет съемный чехол, что существенно облегчает уход за изделием. Со временем, возможна замена матраса и чехла.
- Спальное место формируется одним ровным цельным матрасом без стыков.
Рекомендуемые статьи по данной теме:
КАК РАСКЛАДЫВАТЬ:
Снять подушки сиденья, потянуть механизм вверх и на себя, развернуть звенья, при трансформации становится на стальные опоры – диван разложен.
«ТИК-ТАК»
Механизм предназначен для ежедневного использования.![]() Он прочен и прост в эксплуатации. В основании спального места два одинаковых мягких элемента, спинка и сиденье. Максимальная нагрузка на механизм 240 кг. Раскладка и сон по направлению вдоль стены. Данный механизм трансформации используется в следующих диванах: «Тринити», «Морган», Bormio, Gabrisa, «Соренто».
Он прочен и прост в эксплуатации. В основании спального места два одинаковых мягких элемента, спинка и сиденье. Максимальная нагрузка на механизм 240 кг. Раскладка и сон по направлению вдоль стены. Данный механизм трансформации используется в следующих диванах: «Тринити», «Морган», Bormio, Gabrisa, «Соренто».
ПРЕИМУЩЕСТВА:
- Легкость трансформации.
- Не оставляет следов на напольных покрытиях и коврах, так как перемещается на направляющих, совершая движение-шаг в пространстве. Не используются колесики, что исключает трение с поверхностью пола.
- Под сиденьем находится вместительный бельевой ящик.
- Удобное и просторное спальное место.
- Повышенная степень надежности механизма при ежедневном использовании.
- При раскладке не требуется убирать подушки сиденья.
КАК РАСКЛАДЫВАТЬ:
Для того, чтобы разложить диван нужно снять подушки спинки, затем, взявшись за низ передней части дивана, легким движением вверх и вперед выдвинуть сиденье на себя и аккуратно опустить его на пол.![]() На освободившееся место опустить спинку.
На освободившееся место опустить спинку.
«ТЕЛЕСКОП»
Выкатной или выдвижной механизм трансформации. Особенность конструкции позволяет сделать спальное место высотой только 30 см. Рекомендует как «гостевой вариант» спального места. Максимальная нагрузка 180 кг. Раскладка и сон по направлению от стены.
ПРЕИМУЩЕСТВА:
- Простой в эксплуатации механизм.
- В основании спального места два одинаковых мягких элемента, спинка и сиденье.
- Часто в средней части дивана есть бельевой ящик.
- Компактность в сложенном виде и большое спальное место – в разложенном виде.
КАК РАСКЛАДЫВАТЬ:
Конструкция состоит из 3 частей. Для трансформации в положение «кровать» необходимо выкатить вперед переднюю секцию с сиденьем, при этом из спинки выдвигается подголовник, и уложить сиденье-подушку в образовавшуюся нишу.
«HODRY»
Механизм премиум-класса с электроприводом, производства Австрия. Подходит для ежедневного использования. По принципу разложения – это выкатной механизм. Для трансформации диван-кровати в положение «кровать» достаточно нажать одну кнопку на пульте дистанционного управления. Есть ящик для постельных принадлежностей.
Подходит для ежедневного использования. По принципу разложения – это выкатной механизм. Для трансформации диван-кровати в положение «кровать» достаточно нажать одну кнопку на пульте дистанционного управления. Есть ящик для постельных принадлежностей.
«ВПЕРЕД РАСКЛАДНОЙ»
Удобный механизм, который применяется в двухместных диван-кроватях и креслах-кроватях. В основании спального места два одинаковых мягких элемента, спинка и сиденье. Максимальная нагрузка 200 кг. Есть бельевой ящик. Подходит для ежедневного использования.
КАК РАСКЛАДЫВАТЬ:
- Взявшись за тканевую ручку, закрепленную внизу сидения дивана или в нижней части спинки, поднять передний мягкий элемент спинки вверх, а затем установить его вертикально.
- Поднять вверх сиденье дивана за его переднюю стенку, и перемещая его вперед и на себя, поставить опорами на пол. При этом открывается доступ к вместительному ящику для постельных принадлежностей.
«ПУМА»
Механизм «Пума» предназначен для ежедневного использования и является представителем нового, современного типа механизмов трансформации. Механизм состоит из металлических пружин и противовесов, компенсирующих возможные усилия при раскладке дивана. Прост и надежен в использовании. Бельевого ящика нет. Данный механизм трансформации используется в следующих диванах: «Сиэтл».
ПРЕИМУЩЕСТВА:
- Потрясающая легкость трансформации.
- Не оставляет следов на напольных покрытиях и коврах, так как перемещается на направляющих, совершая поступательное движение-шаг в пространстве. Не используются колесики, что исключает трение с поверхностью пола.
- Комфортное спальное место, сложившееся из двух мягких элементов.
- При раскладке не требуется убирать подушки сиденья.
КАК РАСКЛАДЫВАТЬ:
Раскладка осуществляется путем перемещения сиденья вперед «шагающим» способом.![]() Возьмитесь за специальную петлю подъема (если она есть) или за низ дивана и приподнимите его. В первые же секунды механизм возьмет всю нагрузку на себя и при минимальном усилии с вашей стороны сиденье переместится и сделает шаг вперед. Одновременно с перемещением сиденья вперед автоматически осуществляется подъем мягкого элемента, находящегося в нише под сиденьем. Сиденье и дополнительный мягкий элемент образуют единое спальное место.
Возьмитесь за специальную петлю подъема (если она есть) или за низ дивана и приподнимите его. В первые же секунды механизм возьмет всю нагрузку на себя и при минимальном усилии с вашей стороны сиденье переместится и сделает шаг вперед. Одновременно с перемещением сиденья вперед автоматически осуществляется подъем мягкого элемента, находящегося в нише под сиденьем. Сиденье и дополнительный мягкий элемент образуют единое спальное место.
«ДЕЛЬФИН»
Популярный и удобный механизм для ежедневного применения. Чаще всего, используется в угловых диванах. В основании спального места: сиденье дивана и выдвижной блок, расположенный под сиденьем. Максимальная нагрузка 200 кг. Прочный и надежный механизм. Данный механизм трансформации используется в следующих диванах:«Престон», «Лаунж-2», «Стив».
ПРЕИМУЩЕСТВА:
- В угловых диванах просторное спальное место.
- Легкость трансформации, практически не требуется усилий при раскладывании.
- Надежный стальной механизм, направляющие из твердых пород древесины продлевают срок службы конструкции.
- Не требует дополнительного пространства, раскладка в границах общих габаритов дивана.
- Легкость раскладки позволяет в считанные секунды превратить диван в большую зону для отдыха для нескольких человек. Вы можете расположиться на нем с комфортом, вытянув ноги и облокотившись на подушки спинки.
- При раскладке не требуется убирать подушки сиденья.

КАК РАСКЛАДЫВАТЬ:
Выдвинуть на себя встроенный потайной ящик из-под сиденья дивана. Потянув за ремешок, вытянуть дополнительную секцию, чтобы она сравнялась с высотой сиденья и «достроила» спальное место до полноценного размера.
«ЕВРОКНИЖКА»
«Еврокнижка» — один из самых популярных и востребованных механизмов для ежедневного сна. Очень прост и надежен в использовании. Имеет направляющие из твердых пород древесины или фанеры, по которым передвигаются ролики, установленные сзади внизу сиденья.![]() Передняя часть сиденья также опирается колесными опорами на пол. Под сиденьем всегда имеется вместительный бельевой ящик. Максимальная нагрузка на механизм 240 кг. Раскладка и сон по направлению вдоль стены. Данный механизм трансформации используется в следующих диванах:«Толедо».
Передняя часть сиденья также опирается колесными опорами на пол. Под сиденьем всегда имеется вместительный бельевой ящик. Максимальная нагрузка на механизм 240 кг. Раскладка и сон по направлению вдоль стены. Данный механизм трансформации используется в следующих диванах:«Толедо».
ПРЕИМУЩЕСТВА:
- Максимально простая конструкция механизма гарантирует повышенную надежность при ежедневной трансформации.
- Легкость трансформации.
- При раскладке не требуется убирать подушки сиденья.
- В основании спального места два одинаковых мягких элемента: спинка и сиденье.
КАК РАСКЛАДЫВАТЬ:
Для того, чтобы разложить механизм: снять подушки спинки, сиденье вытягивается на себя, на освободившееся место опускается спинка. Готово.
«РИБАЛТО»
Mеханизм двойного сложения «Рибалто» — спальное место, состоящее из трех частей, две из которых являются до трансформации основанием сидения, а третья, складываясь, образует спинку. Механизм трансформации «Рибалто» обеспечивает легкость и оперативность раскладывания. Диваны с механизмом «Рибалто» в собранном виде занимают минимальную площадь, а при раскладывании имеют широкое и высокое спальное место с комфортным ровным матрацем.
Механизм трансформации «Рибалто» обеспечивает легкость и оперативность раскладывания. Диваны с механизмом «Рибалто» в собранном виде занимают минимальную площадь, а при раскладывании имеют широкое и высокое спальное место с комфортным ровным матрацем.
«АККОРДЕОН»
Один из самых востребованных механизмов для гостевого использования. Работает по принципу гармошки, что обеспечивает легкость и оперативность раскладывания. Диваны с механизмом аккордеон в собранном виде занимают минимальную площадь, а при раскладывании имеют широкое и высокое спальное место. Раскладка и сон по направлению от стены. Рекомендуемая нагрузка 160 кг.
ПРЕИМУЩЕСТВА:
- Экономия пространства.
- Для раскладки не требуется дополнительно убирать никакие элементы с дивана.
- Раскладка одним движением.
- Легкая и простая раскладка.
- Повышенная степень надежности механизма при ежедневном использовании (срок 10 лет).
- Ровное и просторное спальное место без стыков.
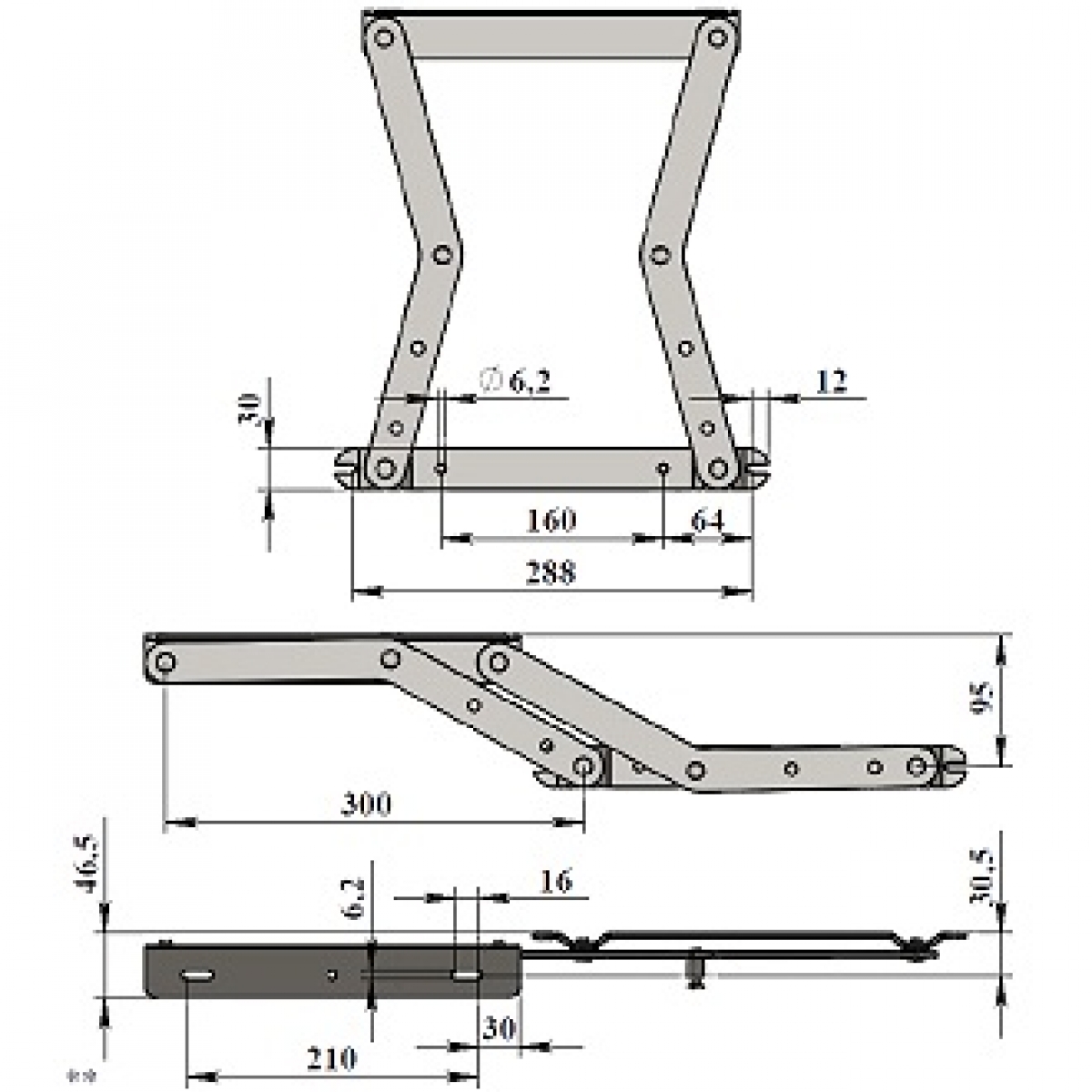
КАК РАСКЛАДЫВАТЬ:
Потяните сиденье дивана вперед, используя специальную петлю или просто схватившись за нижнюю часть изделия. Тяните до тех пор, пока диван полностью не разложится в кровать.
«КНИЖКА»
Для трансформации диван-кровати в положение кровать приподнимается сидение, при этом спинка дивана опускается в горизонтальное положение. Сиденье поднимается далее, до характерного щелчка, после этого опускается также в горизонтальное положение. Для механизма «книжка» необходимо наличие небольшого пространства между спинкой и стеной.Под сиденьем находится вместительный бельевой ящик. Спальное место располагается вдоль стены (важно для владельцев малогабаритных квартир).
«ПУМА ВЫКАТНОЙ»
Интересная разновидность механизма «Пума», которая устанавливается в компактные диваны (длина 210-220) и дает возможность иметь полноценное спальное место для ежедневного сна. Одна из последних разработок в механизмах трансформации, производство — Польша. Механизм основан на принципе синхронизации движений, прост и надежен в использовании. Бельевого ящика нет.
Одна из последних разработок в механизмах трансформации, производство — Польша. Механизм основан на принципе синхронизации движений, прост и надежен в использовании. Бельевого ящика нет.
ПРЕИМУЩЕСТВА:
- Потрясающая легкость трансформации.
- При раскладке не требуется убирать подушки сиденья.
- Позволяет иметь полноценное и долговечное спальное место в компактных диванах.
Чтобы механизм трансформации служил долго, необходимо соблюдать следующие правила эксплуатации:
- В сложенном механизме может находиться только матрас, поставленный в комплекте изделия. В сложенном механизме не допускается присутствие спальных принадлежностей и посторонних предметов.
- При трансформации механизма не следует допускать попадания постельных принадлежностей, посторонних предметов и пальцев между подвижными элементами механизма.
- В случае затрудненного раскладывания или складывания проверьте отсутствие посторонних предметов в шарнирных соединениях, не прилагая излишних усилий.
- Не рекомендуется садиться или опираться на края механизма, не имеющие опоры: подголовник, ножную секцию.
«NEW ROLLER»
Механизм трансформации «New Roller» уникален среди своих собратьев: диван раскладывается с изголовья, благодаря этому подушки снимать не надо — они уходят под низ дивана. Разработан Бельгийской компанией Sedak-meral и, также как и все остальные механизмы этой компании, имеет высокое качество сборки и используемого материала.
ПРЕИМУЩЕСТВА:
- Не встраивается в диван и по сути является основой дивана или его каркасом.
- Не имеет аналогов в простоте сложения-раскладки.
- Возможность прикрутить спинку дивана на дому у покупателя, делает диван «проходимым» в любые дверные проемы.
- Данный механизм выдерживает нагрузку до 500 кг.
- Можно использовать матрас толщиной до 140 мм, что значительно расширяет количество допустимых матрасов к этой модели механизма.
- Каркас выполнен из высококачественной стали толщиной стенок 3 мм.
- Стальная решётка из которой выполнено спальное место занимает 2/3 его поверхности и обеспечивает надёжное и удобное спальное место даже для людей с проблемами позвоночника.

«ЛУКА D30»
В качестве спального места используется механизм трансформации Российского производства «Лука D30» двойного сложения.
Глубина в сложенном состоянии 780 мм. Спальная поверхность в местах наибольшей нагрузки выполнена из сварной сетки, диаметр прутка 4 мм. В ножной части эластичные ремни. Ремни так же позволяют получить более комфортное сидячее место в положении диван. Нагрузка воспринимается не только подушкой дивана, но и матрацем механизма. Тяги выполнены из 5 мм стальной полосы. Данное решение позволяет добиться наибольшей надежности и долговечности работы данного соединения. Головная часть выполнена как единый силовой элемент, состоящий из: П- образной дуги, поперечной трубы диаметром 30 мм с толщиной стенки 1,5 мм, связанные между собой сварной сеткой.![]()
ОСОБЕННОСТИ:
- Данный механизм в разложенном виде позволяет получить спальное место размером 1620 х 1890 мм.
- Высота механизма в разложенном виде до верхнего края матраса составляет 410 мм.
«РАНУЧЧИ»
Итальянцы, законодатели мебельной моды, уверены: сидеть и спать надо на разных элементах дивана, т.к. для сидения требуется более мягкий наполнитель, а для здорового сна – более твёрдый. Механизм двойного сложения «Рануччи» — это уникальный механизм для ежедневного сна, комфорт которого сравним с полноценной кроватью. Спальное место состоит из трех частей, две из которых являются до трансформации основанием сиденья, а третья, складываясь, образует спинку. Диваны с механизмом «Рануччи» в собранном виде занимают минимальную площадь, т.к. сам механизм «Рануччи» является и диваном и спальным местом одновременно, что позволяет экономить пространство в жилом помещении.
Механизм раскладывается за одно движение, подушки сидения и спинки не надо снимать, т. к. они уходят под спальное место. В результате раскладывания мы получаем отличное спальное место, без стыков, без перепадов уровня (что часто бывает, когда используются различные выкатные системы). Матрац сделан из из качественного пенополиуретана, и опорой для него является сварная стальная сетка, закреплённая на раме механизма с помощью пружин (это создаёт дополнительный комфорт).
к. они уходят под спальное место. В результате раскладывания мы получаем отличное спальное место, без стыков, без перепадов уровня (что часто бывает, когда используются различные выкатные системы). Матрац сделан из из качественного пенополиуретана, и опорой для него является сварная стальная сетка, закреплённая на раме механизма с помощью пружин (это создаёт дополнительный комфорт).
Рама механизма сделана из стальных труб, а детали само го механизма – из стального листа. Простоту, комфорт и надежность механизма «Рануччи» по достоинству оценила и выбрала для своей мебели такая всемирно известная сеть отелей, как «Hilton» («Хилтон»).
Механизмы трансформации диванов: как выбрать подходящий?
Механизмы трансформации диванов: как выбрать подходящий?
13 июня 2021
Чаще всего новый диван выбирают по внешнему виду — подходящий под интерьер и с красивой обивкой. На самом деле не меньшее значение имеет механизм раскладывания: именно от него зависит, насколько долго вам прослужит мягкая мебель и удобно ли будет её использовать.![]() Давайте разберемся, как подобрать подходящий механизм трансформации.
Давайте разберемся, как подобрать подходящий механизм трансформации.
Типы диванов по механизму раскладывания
Диваны с механизмом бывают двух видов: гостевые и для ежедневного сна.
Они отличаются по прочности, удобству сна и тому, насколько легко из раскладывать. При выборе учтите, как часто вам придется его раскладывать, какой нужен размер, будете ли вы использовать его для сна. Дальше — изучите достоинства разных механизмов трансформации и выберите наиболее подходящий для вас.
Для ежедневного использования
Если вы подбираете диван именно для сна, обратите внимание на модели с прочным механизмом раскладывания и ортопедическими свойствами. Это могут быть «еврокнижка», «тик-так», «пума», «остин», «высоковыкатной», «дельфин» или «кушетка». Надежные и прочные, такие механизмы прослужат долго даже при постоянном использовании.
На диване для повседневного сна должно быть удобно, иначе вы рискуете проснуться с болью в спине. Поверхность перечисленных выше диванов такая же комфортная, как ортопедический матрас, поэтому они подходят для ежедневного сна. Они незаменимы в спальнях и детских комнатах — днем экономят место, а ночью на них комфортно спать.
Они незаменимы в спальнях и детских комнатах — днем экономят место, а ночью на них комфортно спать.
Многие раскладные диваны заменяют двуспальную кровать, другие — рассчитаны на одного человека. Так что подходящий трансформер можно подобрать и для семейной пары, и для ребенка, и для холостяка.
Гостевые
Диваны с гостевыми видами механизмов не так удобно трансформировать, как с механизмами для ежедневного использования. Чаще всего гостевые диваны рассчитаны на мягкую и комфортную посадку, а спальное место скорее как бонус.
К таким механизмам относятся: «французская раскладушка», «франко-бельгийская раскладушка» и «итальянская раскладушка», которую еще называют «спартак». Диваны такого типа незаменимы, когда нужно организовать временные спальные места, например, в случае приезда гостей.
Виды механизмов трансформации
Механизмы раскладки диванов постоянно модернизируются. У обычной «книжки» уже есть несколько улучшенных вариантов: «еврокнижка», «клик-кляк», «пантограф».
Среди новых видов трансформации диванов — механизм «реклайнер», который встраивают, в основном, в кресла. Это не полноценное спальное место, но на нем приятно вздремнуть или расслабиться за просмотром фильма. Рассмотрим подробно самые популярные виды.
Еврософа (Еврокнижка)
«Еврософу» сможет разложить даже ребенок, поэтому они так популярны. Из-за минимального количества элементов такой механизм редко ломается. Механизм раскладывания устроен максимально просто: вам надо потянуть сиденье вперед, а на его место поставить спинку. Поверхность получается ровной и большой по площади, а во внутренних ящиках можно хранить постельное белье, подушки и одеяла.
Преимущества
— прочный;
— просторный;
— в собранном виде — полноценное спальное место на одного человека;
— легко раскладывается;
— имеет встроенный отсек для белья.
Тик-Так (Пантограф)
Диваны с механизмом «Тик-Так» называют также «шагающей еврокнижкой» из-за похожего принципа раскладывания. Главная его особенность в том, что при раскладке сиденье не выкатывается на роликах, а перешагивает нужное расстояние, не задевая линолеум, ламинат и даже ковер.
Главная его особенность в том, что при раскладке сиденье не выкатывается на роликах, а перешагивает нужное расстояние, не задевая линолеум, ламинат и даже ковер.
«Тик-Так» легко раскладывается: вам надо приподнять сиденье и потянуть его вперед, а на освободившееся мест вложить спинку. Спать на нем так же удобно, как и на «еврокнижке».
Преимущества:
— просторный;
— в собранном виде — полноценное спальное место на одного человека;
— имеет встроенный отсек для белья;
— легко трансформируется.
Дельфин
Механизм получил название из-за особого складывания: спальное место задвигается под сиденье дивана — визуально это напоминает нырок дельфина в воду. Некоторые мебельные фабрики называют называют этот механизм «кенгуру», нетрудно догадаться почему.
Диваны с механизмом «дельфин» раскладывать просто: нужно выдвинуть нижнюю часть дивана из-под сиденья, а затем потянуть ее вверх за ремешок, пока подушки не сравняются. «Дельфин» часто устанавливают на угловые модели, в таком случае отсек для белья находится в неподвижной части — угловом модуле (канапе).
«Дельфин» часто устанавливают на угловые модели, в таком случае отсек для белья находится в неподвижной части — угловом модуле (канапе).
Преимущества:
— просторный;
— в разобранном виде не занимает много места;
— надежный;
— легко трансформируется.
Пума / Остин
Механизм «пума» по типу трансформации напоминает «тик-так». Чтобы его разложить, нужно поднять сиденье и выдвинуть его вперед, а подушку под сиденьем поднять наверх, чтобы обе части сравнялись и образовали ровное спальное место. Благодаря пружинам в механизме, часть трансформации происходит автоматически, поэтому особых усилий прикладывать не нужно.
У «пумы» нет выдвижных роликов, поэтому они не оставят вмятин на линолеуме и не поцарапают ламинат.
Еще один похожий на «пуму» механизм — «остин». Он раскладывается по такому же принципу, но сделан из трех частей: две части поднимаются и раскладываются, а третья вытягивается из-под сиденья.
Преимущества:
— компактный;
— надежный;
— удобен для сна.![]()
Высоковыкатной
«Высоковыкатной» напоминает сразу два механизма раскладки диванов: «выкатной» и «дельфин». Он состоит из нескольких частей и выдвигается вперед, как выкатные модели, но одна из деталей разбирается по сходному принципу с «дельфином» — вытягивается и поднимается вверх.
«Высоковыкатной» перенял лучшие качества двух механизмов сразу: спать на нем также высоко и комфортно, как на моделях с «дельфином», а в собранном виде выглядит компактно и экономит место в комнате. Благодаря длинной и ровной поверхности, спать на нем комфортно, как на двуспальной кровати.
Преимущества:
— большая площадь;
— надежный;
— высокое и ровное спальное место;
— подойдет для небольших помещений.
Кушетка
«Кушеткой» называют небольшие диваны, которые разбираются в одноместную кровать, но раскладываться они могут по-разному.
Встречаются два разных механизма под названием «кушетка». Первый трансформируется в кровать, благодаря мягким подлокотникам, которые опускаются и образуют ровную вытянутую поверхность.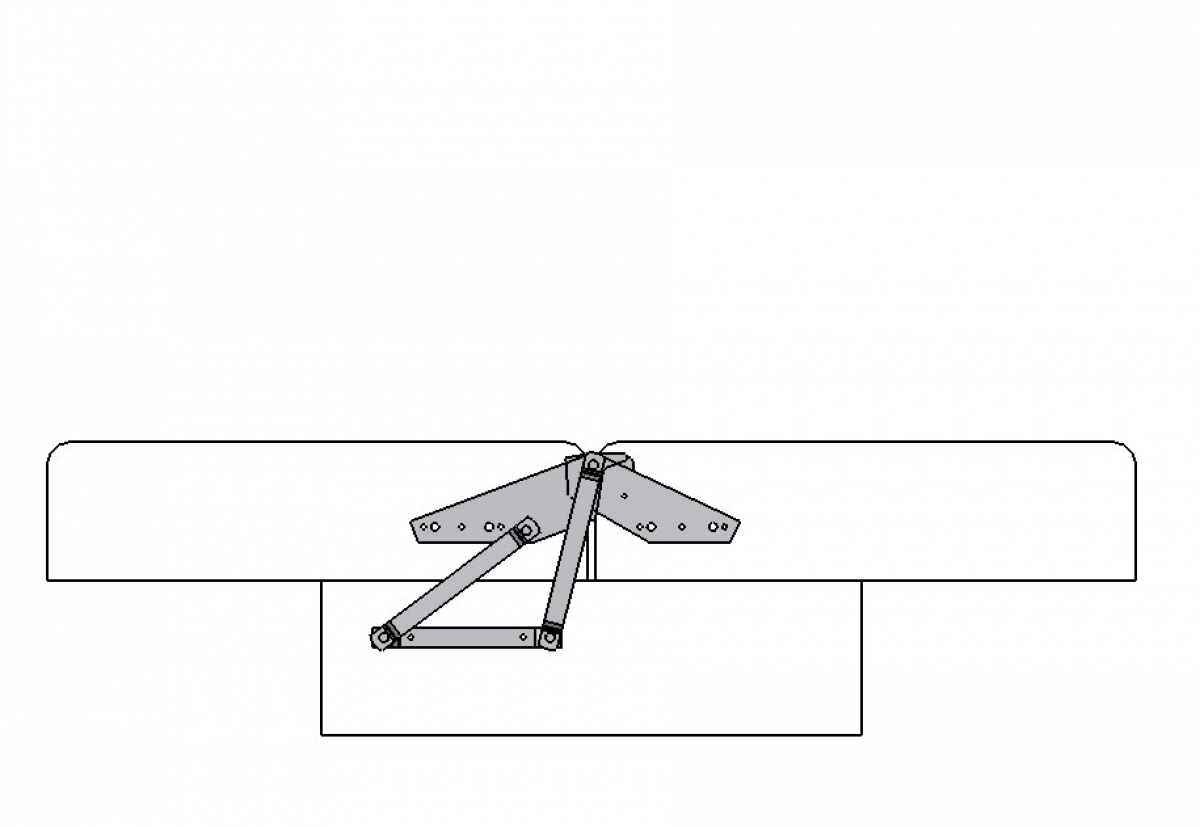 Второй — похож на механизм раскладки «еврокнижка». Его боковая часть выдвигается и оставляет место под изголовье дивана, которое туда опускается.
Второй — похож на механизм раскладки «еврокнижка». Его боковая часть выдвигается и оставляет место под изголовье дивана, которое туда опускается.
Даже после трансформации «кушетка» занимает мало места. Идеально подойдет для детских комнат, спален для одного человека и даже лоджий, где иногда так хочется вздремнуть.
Преимущества:
— легко трансформируется в кровать;
— есть отсек для белья;
— вариант для небольшой комнаты.
Французская раскладушка (Миксотойл)
Диваны с механизмом «французская раскладушка» лучше использовать для сидения, чем для сна. Рекомендуем использовать механизм как гостевой вариант спального места.
Сама раскладушка находится под сиденьем. Чтобы ее разобрать, нужно убрать подушки и потянуть за край сиденья. Части раскладушки раздвинутся и встанут на ножки.
Преимущества:
— просторный;
— компактный.
Франко-бельгийская раскладушка (Седафлекс)
Он одновременно прост в применении и надежен, что позволяет считать «седафлекс» идеальным вариантом трансформации для более частого использования. Спальное место получается идеально ровным, без изгибов и заломов, а сам раскладной механизм создается из высококачественного металла и потому служит долго.
Спальное место получается идеально ровным, без изгибов и заломов, а сам раскладной механизм создается из высококачественного металла и потому служит долго.
Усиленный металлический каркас, металлическая сетка, возможность выбора 12- сантиметрового матраса (ППУ или пружинного), жесткость конструкции- все это обеспечивает комфортный и спокойный сон. Механизм выдерживает нагрузку в 150 кг. на каждое спальное место.
Большим достоинством «седафлекса» является также и то, что в сложенном виде он имеет параметры, позволяющие сделать настил сиденья максимально удобным.
Преимущества:
— просторный;
— компактный;
— надежный;
— матрас на выбор ППУ или ПБ (пружинный блок).
Подводим итоги: диван с каким механизмом лучше
Идеального механизма трансформации не существует: каждый из них отвечает определенным требованиям. Определите, чего именно вы ждете от нового дивана, и отталкивайтесь от этого при выборе модели. Так вы сможете подобрать раскладной диван, который прослужит вам долгие годы.
Бактериальная трансформация: распространение, общие механизмы и дивергентный контроль
Гриффит Ф. Значение типов пневмококка. J. Hyg. 27 , 113–159 (1928).
CAS PubMed Google Scholar
Краучер, Н. Дж. И др. Быстрая эволюция пневмококка в ответ на клиническое вмешательство. Наука 331 , 430–434 (2011).
CAS PubMed PubMed Central Google Scholar
Лоренц, М.G. & Wackernagel, W. Бактериальный перенос генов путем естественной генетической трансформации в окружающей среде. Microbiol. Ред. 58 , 563–602 (1994).
CAS PubMed PubMed Central Google Scholar
Джонсборг О.![]() , Элдхольм В. и Ховарштейн Л. С. Естественная генетическая трансформация: распространенность, механизмы и функции. Res. Microbiol. 158 , 767–778 (2007).
, Элдхольм В. и Ховарштейн Л. С. Естественная генетическая трансформация: распространенность, механизмы и функции. Res. Microbiol. 158 , 767–778 (2007).
CAS PubMed Google Scholar
Аверхофф, Б.& Фридрих, А. Системы естественной трансформации, связанные с пилями IV типа: транспорт ДНК в мезофильных и термофильных бактериях. Arch. Microbiol. 180 , 385–393 (2003).
CAS PubMed Google Scholar
Чен И. и Дубнау Д. Поглощение ДНК во время бактериальной трансформации. Nature Rev. Microbiol. 2 , 241–249 (2004).
CAS Google Scholar
Аллеманд, Дж.Ф. и Майер Б. Моторы бактериальной транслокации исследуются с помощью методов одиночных молекул. FEMS Microbiol. Ред. 33 , 593–610 (2009).
CAS PubMed Google Scholar
Клаверис, Дж. П., Мартин, Б. и Полард, П. Механизм генетической трансформации: состав, локализация и механизм. FEMS Microbiol. Ред. 33 , 643–656 (2009).
П., Мартин, Б. и Полард, П. Механизм генетической трансформации: состав, локализация и механизм. FEMS Microbiol. Ред. 33 , 643–656 (2009).
CAS PubMed Google Scholar
Аллеманд, Дж.Ф., Майер Б. и Смит Д. Е. Молекулярные моторы для транслокации ДНК у прокариот. Curr. Opin. Biotechnol. 23 , 503–509 (2012).
CAS PubMed PubMed Central Google Scholar
Клаверис, Дж. П., Прюдом, М. и Мартин, Б. Индукция регулонов компетенции как общие реакции на стресс у грамположительных бактерий. Annu. Rev. Microbiol. 60 , 451–475 (2006).
CAS PubMed Google Scholar
Зейтц, П. и Блокеш, М. Сигналы и регуляторные пути, участвующие в естественной компетентности и трансформации патогенных и экологических грамотрицательных бактерий. FEMS Microbiol. Ред. 37 , 336–363 (2012).
Ред. 37 , 336–363 (2012).
PubMed Google Scholar
Драшкович И. и Дубнау Д. Биогенез предполагаемого канального белка ComEC, необходимого для поглощения ДНК: топология мембраны, олигомеризация и образование дисульфидных связей. Мол. Microbiol. 55 , 881–896 (2005).
CAS PubMed Google Scholar
Mortier-Barrière, I. et al. Ключевая пресинаптическая роль в трансформации широко распространенного бактериального белка: DprA передает входящую оцДНК к RecA. Cell 130 , 824–836 (2007). Это исследование идентифицирует DprA как предназначенный для трансформации загрузчик RecA на трансформирующую оцДНК, что является важным этапом для образования рекомбинантов трансформации.
PubMed Google Scholar
Хоббс, М. и Маттик, Дж. С. Общие компоненты в сборке фимбрий типа 4, системы переноса ДНК, нитчатый фаг и аппарат для секреции белков: общая система для образования поверхностно-ассоциированных белковых комплексов. Мол. Microbiol. 10 , 233–243 (1993).
Мол. Microbiol. 10 , 233–243 (1993).
CAS PubMed Google Scholar
Дубнау, Д.Поглощение ДНК бактериями. Annu. Rev. Microbiol. 53 , 217–244 (1999).
CAS PubMed PubMed Central Google Scholar
Манн, Дж. М., Карабетта, В. Дж., Кристя, И. М. и Дубнау, Д. Комплексное образование и переработка пилинов минорной трансформации Bacillus subtilis . Мол. Microbiol. 90 , 1201–1215 (2013).
CAS PubMed PubMed Central Google Scholar
Чен, И., Provvedi, R. & Dubnau, D. Макромолекулярный комплекс, образованный пилин-подобным белком в компетентных Bacillus subtilis . J. Biol. Chem. 281 , 21720–21727 (2006). В этой статье документально подтверждено наличие короткой пилюсовидной структуры у B. subtilis . Это первая идентификация специфического псевдопилуса у грамположительных бактерий.
subtilis . Это первая идентификация специфического псевдопилуса у грамположительных бактерий.
CAS PubMed Google Scholar
Laurenceau, R.и другие. Пилус типа IV опосредует связывание ДНК во время естественной трансформации в Streptococcus pneumoniae . PLoS Pathog. 9 , e1003473 (2013). Путем идентификации длинных пилусов, которые необходимы для трансформации у S. pneumoniae , это исследование показывает, что трансформационные пили могут выходить далеко за пределы клеточной стенки грамположительных бактерий и захватывать экзогенную ДНК.
CAS PubMed PubMed Central Google Scholar
Провведи, Р.& Dubnau, D. ComEA представляет собой рецептор ДНК для трансформации компетентной Bacillus subtilis . Мол. Microbiol. 31 , 271–280 (1999).
CAS PubMed Google Scholar
Берроуз, Л. Л. Оружие массового втягивания. Мол. Microbiol. 57 , 878–888 (2005).
Л. Оружие массового втягивания. Мол. Microbiol. 57 , 878–888 (2005).
CAS PubMed Google Scholar
Пуйе, А., Гринберг, Б. и Лакс, С. А. Генетическая и структурная характеристика EndA. Мембраносвязанная нуклеаза, необходимая для трансформации Streptoccus pneumoniae . J. Mol. Биол. 213 , 727–738 (1990).
CAS PubMed Google Scholar
Bergé, M. et al. Поглощение трансформирующей ДНК у грамположительных бактерий: взгляд из Streptococcus pneumoniae . Мол. Microbiol. 45 , 411–421 (2002).
PubMed Google Scholar
Зейтц, П. и Блокеш, М. Механизм захвата ДНК естественно компетентных Vibrio cholerae . Proc. Natl Acad. Sci. США 110 , 17987–17992 (2013).
CAS Google Scholar
Bergé, M. J. et al. Вовлечение в средние клетки нуклеазы поглощения ДНК и вирулентности, EndA, для трансформации пневмококка. PLoS Pathog. 9 , e1003596 (2013). Это исследование показывает, что нуклеаза EndA рекрутируется в среднюю клетку во время пневмококковой компетентности, что указывает на то, что поглощение ДНК, вероятно, происходит в этом месте.
J. et al. Вовлечение в средние клетки нуклеазы поглощения ДНК и вирулентности, EndA, для трансформации пневмококка. PLoS Pathog. 9 , e1003596 (2013). Это исследование показывает, что нуклеаза EndA рекрутируется в среднюю клетку во время пневмококковой компетентности, что указывает на то, что поглощение ДНК, вероятно, происходит в этом месте.
PubMed PubMed Central Google Scholar
Hahn, J. et al. Белки трансформации и захват ДНК локализуются на полюсах клетки в Bacillus subtilis . Cell 122 , 59–71 (2005). Это исследование показывает, что захват ДНК происходит на полюсах клеток у компетентных B. subtilis .
CAS PubMed PubMed Central Google Scholar
Лондоньо-Вальехо, Дж. А. и Дубнау, Д. Мутация предполагаемого сайта связывания нуклеотидов мембранного белка Bacillus subtilis ComFA отменяет захват ДНК во время трансформации. J. Bacteriol. 176 , 4642–4645 (1994).
J. Bacteriol. 176 , 4642–4645 (1994).
PubMed PubMed Central Google Scholar
Yeh, Y. C., Lin, T. L., Chang, K. C. & Wang, J. T. Характеристика гомолога ComE3, необходимого для трансформации ДНК в Helicobacter pylori . Заражение. Иммун. 71 , 5427–5431 (2003).
CAS PubMed PubMed Central Google Scholar
Стингл, К.и другие. Композитная система опосредует двухэтапный захват ДНК в Helicobacter pylori . Proc. Natl Acad. Sci. США 107 , 1184–1189 (2010).
PubMed Google Scholar
Beernink, H. T. & Morrical, S. W. RMP: белки-медиаторы рекомбинации / репликации. Trends Biochem. Sci. 24 , 385–389 (1999).
CAS PubMed Google Scholar
Quevillon-Cheruel, S. и другие. Структурно-функциональный анализ пневмококкового белка DprA показывает, что димеризация имеет решающее значение для загрузки рекомбиназы RecA на ДНК во время трансформации. Proc. Natl Acad. Sci. США 109 , E2466 – E2475 (2012).
и другие. Структурно-функциональный анализ пневмококкового белка DprA показывает, что димеризация имеет решающее значение для загрузки рекомбиназы RecA на ДНК во время трансформации. Proc. Natl Acad. Sci. США 109 , E2466 – E2475 (2012).
CAS PubMed Google Scholar
Prudhomme, M. et al. Стресс антибиотиками вызывает генетическую трансформируемость человеческого патогена Streptococcus pneumoniae . Science 313 , 89–92 (2006). В этой статье сообщается об открытии того, что пневмококковая компетентность индуцируется в ответ на стрессы окружающей среды, такие как антибиотики и агенты, повреждающие ДНК.
CAS PubMed Google Scholar
Шарпантье, X., Полард, П. и Клаверис, Дж. П. Индукция способности к генетической трансформации антибиотиками: конвергентная эволюция стрессовых реакций у отдаленных видов бактерий, лишенных SOS? Curr. Opin.Microbiol. 15 , 1–7 (2012).
Opin.Microbiol. 15 , 1–7 (2012).
Google Scholar
Бутала М., Згур-Берток Д. и Басби С. Дж. Бактериальный репрессор транскрипции LexA. Ячейка. Мол. Life Sci. 66 , 82–93 (2009).
CAS PubMed Google Scholar
Ли, М. С. и Моррисон, Д. А. Идентификация нового регулятора в Streptococcus pneumoniae , связывающего определение кворума со способностью к генетической трансформации. J. Bacteriol. 181 , 5004–5016 (1999).
CAS PubMed PubMed Central Google Scholar
Peterson, S. et al. Выявление компетентных генов, отвечающих за феромон, у Streptococcus pneumoniae . Мол. Microbiol. 51 , 1051–1070 (2004).
CAS PubMed Google Scholar
Мартин, Б. , Quentin, Y., Fichant, G. & Claverys, J. P. Независимая эволюция каскадов регуляции компетентности у стрептококков? Trends Microbiol. 14 , 339–345 (2006). В этой статье представлен филогенетический анализ стрептококкового ComDE TCS и установлено, что S. mutans и несколько других стрептококков не имеют этого главного регулятора компетентности, что предполагает наличие альтернативной цепи регуляции компетентности у этих видов.
, Quentin, Y., Fichant, G. & Claverys, J. P. Независимая эволюция каскадов регуляции компетентности у стрептококков? Trends Microbiol. 14 , 339–345 (2006). В этой статье представлен филогенетический анализ стрептококкового ComDE TCS и установлено, что S. mutans и несколько других стрептококков не имеют этого главного регулятора компетентности, что предполагает наличие альтернативной цепи регуляции компетентности у этих видов.
CAS PubMed Google Scholar
Fontaine, L. et al. Новая система определения кворума феромонов контролирует развитие естественной компетентности у Streptococcus thermophilus и Streptococcus salivarius . J. Bacteriol. 192 , 1444–1454 (2010).
CAS PubMed Google Scholar
Машберн-Уоррен, Л., Morrison, D. A. & Federle, M. J. Новый феромон двойного триптофанового пептида контролирует компетентность в Streptococcus spp. через регулятор Rgg. Мол. Microbiol. 78 , 589–606 (2010).
через регулятор Rgg. Мол. Microbiol. 78 , 589–606 (2010).
CAS PubMed PubMed Central Google Scholar
Morrison, D. A., Guedon, E. & Renault, P. Компетенция в отношении естественной генетической трансформации Streptococcus bovis , группы Streptococci S.Infantarius и S. macedonicus . J. Bacteriol. 195 , 2612–2620 (2013).
CAS PubMed PubMed Central Google Scholar
Morikawa, K. et al. Экспрессия гена загадочного вторичного сигма-фактора раскрывает естественную способность к трансформации ДНК у Staphylococcus aureus . PLoS Pathog. 8 , e1003003 (2012).
CAS PubMed PubMed Central Google Scholar
Мохан, С.И Дубнау Д. Регуляция транскрипции comC : доказательства наличия фактора транскрипции, специфичного для компетенции Bacillus subtilis . J. Bacteriol. 172 , 4064–4071 (1990).
J. Bacteriol. 172 , 4064–4071 (1990).
CAS PubMed PubMed Central Google Scholar
Маамар, Х. и Дубнау, Д. Бистабильность в системе К-состояний (компетенции) Bacillus subtilis требует наличия петли положительной обратной связи. Мол. Microbiol. 56 , 615–624 (2005). В этой статье показано, что стохастическая вариация экспрессии comK в популяциях B. subtilis определяет, какие клетки становятся компетентными.
CAS PubMed Google Scholar
Smits, W. K. et al. Stripping Bacillus : автостимуляция ComK отвечает за бистабильный ответ в развитии компетентности. Мол. Microbiol. 56 , 604–614 (2005).
CAS PubMed Google Scholar
Дубнау Д. и Лосик Р. Бистабильность у бактерий. Мол. Microbiol. 61 , 564–572 (2006).
Бистабильность у бактерий. Мол. Microbiol. 61 , 564–572 (2006).
CAS PubMed Google Scholar
Смитс, В. К., Койперс, О. П. и Вининг, Дж. У. Фенотипическая изменчивость у бактерий: роль регуляции обратной связи. Nature Rev. Microbiol. 4 , 259–271 (2006).
CAS Google Scholar
Maamar, H., Raj, A. & Dubnau, D. Шум в экспрессии генов определяет судьбу клеток в Bacillus subtilis . Наука 317 , 526–529 (2007).
CAS PubMed Google Scholar
Лейснер, М., Стингл, К., Фрей, Э.И Майер Б. Стохастический переход к компетенции. Curr. Opin. Microbiol. 11 , 553–559 (2008).
CAS PubMed Google Scholar
Берка Р. М. и др. Анализ микроматрицы K-состояния Bacillus subtilis : изменения экспрессии по всему геному в зависимости от ComK. Мол. Microbiol. 43 , 1331–1345 (2002).
Мол. Microbiol. 43 , 1331–1345 (2002).
CAS PubMed Google Scholar
Hamoen, L.W. et al. Повышение прогностической ценности сайта связывания фактора транскрипции компетентности (ComK) в Bacillus subtilis с использованием геномного подхода. Nucl. Acids Res. 202 , 5517–5528 (2002).
Google Scholar
Ogura, M. et al. Полногеномный анализ генов, регулируемых транскрипционным фактором ComK Bacillus subtilis . J. Bacteriol. 184 , 2344–2351 (2002).
CAS PubMed PubMed Central Google Scholar
Кузниар А., Ван Хэм Р. К., Понгор С. и Леуниссен Дж. А. Поиск ортологов: поиск соответствующего гена в геномах. Trends Genet. 24 , 539–551 (2008).
CAS PubMed Google Scholar
Уайз, Э. М. младший, Александр, С. П. и Пауэрс, М.Аденозин 3 ‘: 5’-циклический монофосфат как регулятор бактериальной трансформации. Proc. Natl Acad. Sci. США 70 , 471–474 (1973).
М. младший, Александр, С. П. и Пауэрс, М.Аденозин 3 ‘: 5’-циклический монофосфат как регулятор бактериальной трансформации. Proc. Natl Acad. Sci. США 70 , 471–474 (1973).
CAS PubMed Google Scholar
Дорочич И. Э., Уильямс П. М. и Редфилд Р. Дж. Ген аденилатциклазы Haemophilus influenzae : клонирование, последовательность и важная роль в компетентности. J. Bacteriol. 175 , 7142–7149 (1993).
CAS PubMed PubMed Central Google Scholar
Chandler, M. S. Ген, кодирующий белок рецептора цАМФ, необходим для развития компетентности у Haemophilus influenzae Rd. Proc. Natl Acad. Sci. США 89 , 1626–1630 (1992).
CAS PubMed Google Scholar
Редфилд Р. Дж. sxy-1 , мутация Haemophilus influenzae , вызывающая значительное усиление спонтанной компетентности.![]() J. Bacteriol. 173 , 5612–5618 (1991).
J. Bacteriol. 173 , 5612–5618 (1991).
CAS PubMed PubMed Central Google Scholar
Macfadyen, L.P. Регулирование развития компетентности в Haemophilus influenzae . J. Theor. Биол. 207 , 349–359 (2000).
CAS PubMed Google Scholar
Redfield, R.J. et al.Новый CRP-зависимый регулон контролирует экспрессию генов компетентности в Haemophilus influenzae . J. Mol. Биол. 347 , 735–747 (2005).
CAS PubMed Google Scholar
Meibom, K. L. et al. Хитин вызывает естественную компетентность в Vibrio cholerae . Наука 310 , 1824–1827 (2005). В данной статье сообщается об открытии этой компетенции в области V.cholerae индуцируется хитином, который представляет собой сахарный полимер, который широко распространен в водной среде обитания этой бактерии.
CAS Google Scholar
Blokesch, M. Переключатель, опосредованный восприятием кворума, способствует естественной трансформации Vibrio cholerae . Моб. Genet. Элементы 2 , 224–227 (2012).
PubMed PubMed Central Google Scholar
Ло Скрудато, М.& Blokesch, M. Регулятор транскрипции, связывающий восприятие кворума и индукцию хитина, чтобы сделать Vibrio cholerae естественным образом трансформируемым. Nucleic Acids Res. 41 , 3644–3658 (2013). Эта статья представляет собой первую документацию о двусторонней регуляции генов компетенции, в которой они напрямую контролируются двумя отдельными регуляторами.
CAS PubMed PubMed Central Google Scholar
Hamoen, L.В., Венема, Г. и Койперс, О. П. Управляющая компетенция в Bacillus subtilis : совместное использование регуляторов. Микробиология 149 , 9–17 (2003).
Микробиология 149 , 9–17 (2003).
CAS PubMed Google Scholar
Hamoen, LW, Van Werkhoven, AF, Dubnau, D. & Venema, G. Фактор транскрипции компетентности Bacillus subtilis распознает короткие A / T-богатые последовательности, расположенные в уникальном гибком узоре вдоль ДНК спираль. Genes Dev. 12 , 1539–1550 (1998).
CAS PubMed PubMed Central Google Scholar
Hamoen, L. W., Van Werkhoven, A. F., Venema, G. & Dubnau, D. Регулятор плейотропного ответа DegU функционирует как примирующий белок в развитии компетентности у Bacillus subtilis . Proc. Natl Acad. Sci. США 97 , 9246–9251 (2000).
CAS PubMed Google Scholar
Hamoen, L.W. et al. Регулятор переходного состояния Bacillus subtilis AbrB связывается с промоторной областью -35 comK.![]() FEMS Microbiol. Lett. 218 , 299–304 (2003).
FEMS Microbiol. Lett. 218 , 299–304 (2003).
CAS PubMed Google Scholar
Серрор, П. и Соненшейн, А. Л. CodY необходим для нутритивной репрессии генетической компетентности Bacillus subtilis . J. Bacteriol. 178 , 5910–5915 (1996).
CAS PubMed PubMed Central Google Scholar
Hoa, T. T., Tortosa, P., Albano, M. & Dubnau, D. Rok (YkuW) регулирует генетическую компетентность в Bacillus subtilis путем прямого подавления comK . Мол. Microbiol. 43 , 15–26 (2002).
CAS PubMed Google Scholar
Тургай, К., Хамоэн, Л.W., Venema, G. и Dubnau, D. Биохимическая характеристика молекулярного переключателя с участием белка теплового шока ClpC, который контролирует активность ComK, фактора транскрипции компетентности Bacillus subtilis .![]() Genes Dev. 11 , 119–128 (1997). В этой статье in vitro описывается взаимодействие между ComK, ClpCP, MecA и ComS и показано, что накопление B.subtilis ComK регулируется протеолизом.
Genes Dev. 11 , 119–128 (1997). В этой статье in vitro описывается взаимодействие между ComK, ClpCP, MecA и ComS и показано, что накопление B.subtilis ComK регулируется протеолизом.
CAS PubMed Google Scholar
Turgay, K., Hahn, J., Burghoorn, J. & Dubnau, D. Компетенция в Bacillus subtilis контролируется регулируемым протеолизом фактора транскрипции. EMBO J. 17 , 6730–6738 (1998).
CAS PubMed PubMed Central Google Scholar
Накано, М.M. et al. srfA — это оперон, необходимый для продукции сурфактина, развития компетентности и эффективного споруляции у Bacillus subtilis . J. Bacteriol. 173 , 1770–1778 (1991).
CAS PubMed PubMed Central Google Scholar
Roggiani, M.![]() & Dubnau, D. ComA, фосфорилированный белок-регулятор ответа Bacillus subtilis , связывается с промоторной областью srfA . J. Bacteriol. 175 , 3182–3187 (1993).
& Dubnau, D. ComA, фосфорилированный белок-регулятор ответа Bacillus subtilis , связывается с промоторной областью srfA . J. Bacteriol. 175 , 3182–3187 (1993).
CAS PubMed PubMed Central Google Scholar
Hayashi, K. et al. Чувствительный к стрессу регулятор h3O2 PerR положительно регулирует экспрессию srfA в Bacillus subtilis . J. Bacteriol. 187 , 6659–6667 (2005).
CAS PubMed PubMed Central Google Scholar
Хуэй, Ф.М. и Моррисон, Д. А. Генетическая трансформация в Streptococcus pneumoniae : анализ нуклеотидной последовательности показывает, что comA , ген, необходимый для индукции компетентности, является членом семейства бактериальных АТФ-зависимых транспортных белков. J. Bacteriol. 173 , 372–381 (1991).
CAS PubMed PubMed Central Google Scholar
Пестова, Э. В., Ховарштейн, Л. С. и Моррисон, Д.A. Регулирование способности к генетической трансформации в Streptococcus pneumoniae с помощью аутоиндуцированного пептидного феромона и двухкомпонентной регуляторной системы. Мол. Microbiol. 21 , 853–864 (1996).
В., Ховарштейн, Л. С. и Моррисон, Д.A. Регулирование способности к генетической трансформации в Streptococcus pneumoniae с помощью аутоиндуцированного пептидного феромона и двухкомпонентной регуляторной системы. Мол. Microbiol. 21 , 853–864 (1996).
CAS PubMed Google Scholar
Håvarstein, L. S., Coomaraswamy, G. & Morrison, D. A. Немодифицированный феромон гептадекапептида индуцирует способность к генетической трансформации у Streptococcus pneumoniae . Proc. Natl Acad. Sci. США 92 , 11140–11144 (1995).
PubMed Google Scholar
Martin, B. et al. Взаимодействие ComE / ComE∼P диктует статус активации или исчезновения пневмококкового X-состояния (компетентности). Мол. Microbiol. 87 , 394–411 (2012).
PubMed Google Scholar
Gardan, R.![]() et al. Система транспорта олигопептидов важна для развития естественной компетентности у штамма Streptococcus thermophilus LMD-9. J. Bacteriol. 191 , 4647–4655 (2009). В этой статье сообщается об основополагающем открытии участия переносчика олигопептидов в регуляторном каскаде компетенции, которое проложило путь к открытию ComRS у S. thermophilus .
et al. Система транспорта олигопептидов важна для развития естественной компетентности у штамма Streptococcus thermophilus LMD-9. J. Bacteriol. 191 , 4647–4655 (2009). В этой статье сообщается об основополагающем открытии участия переносчика олигопептидов в регуляторном каскаде компетенции, которое проложило путь к открытию ComRS у S. thermophilus .
CAS PubMed PubMed Central Google Scholar
Gardan, R. et al. Внеклеточный жизненный цикл ComS, пептида, стимулирующего компетенцию Streptococcus thermophilus . J. Bacteriol. 195 , 1845–1855 (2013).
CAS PubMed PubMed Central Google Scholar
Fontaine, L. et al. Механизм активации компетентности сигнальной системой ComRS у стрептококков. Мол. Microbiol. 87 , 1113–1132 (2013).
CAS PubMed Google Scholar
Сулавик, М. К., Tardif, G. & Clewell, D. B. Идентификация гена rgg , который регулирует экспрессию глюкозилтрансферазы и влияет на фенотип Spp Streptococcus gordonii Challis. J. Bacteriol. 174 , 3577–3586 (1992).
К., Tardif, G. & Clewell, D. B. Идентификация гена rgg , который регулирует экспрессию глюкозилтрансферазы и влияет на фенотип Spp Streptococcus gordonii Challis. J. Bacteriol. 174 , 3577–3586 (1992).
CAS PubMed PubMed Central Google Scholar
Fleuchot, B. et al. Белки Rgg, связанные с интернализованными небольшими гидрофобными пептидами: новый механизм определения кворума у стрептококков. Мол. Microbiol. 80 , 1102–1119 (2011).
CAS PubMed Google Scholar
de Saizieu, A. et al. Идентификация на основе микрочипов нового регулона Streptococcus pneumoniae , контролируемого аутоиндуцированным пептидом. J. Bacteriol. 182 , 4696–4703 (2000).
CAS PubMed PubMed Central Google Scholar
Федерле, М. J. & Morrison, D. A. Один, если по суше, два, если по морю: сигнализация рядов с CSP и XIP. Мол. Microbiol. 86 , 241–245 (2012).
J. & Morrison, D. A. Один, если по суше, два, если по морю: сигнализация рядов с CSP и XIP. Мол. Microbiol. 86 , 241–245 (2012).
CAS PubMed PubMed Central Google Scholar
Lemme, A. et al. Субпопуляционный анализ транскриптома индуцированного пептидом, стимулирующего компетентность, Streptococcus mutans . J. Bacteriol. 193 , 1863–1877 (2011).
CAS PubMed PubMed Central Google Scholar
Son, M. et al. Микрожидкостное исследование регуляции компетентности у Streptococcus mutans : входы окружающей среды модулируют бимодальную и унимодальную экспрессию comX . Мол. Microbiol. 86 , 258–272 (2012).
CAS PubMed PubMed Central Google Scholar
Мироуз, Н.и другие. Прямое участие DprA, загрузчика RecA, предназначенного для трансформации, в отключении пневмококковой инфекции. Proc. Natl Acad. Sci. США 110 , E1035 – E1044 (2013).
Proc. Natl Acad. Sci. США 110 , E1035 – E1044 (2013).
CAS PubMed Google Scholar
Ямамото, С., Морита, М., Изумия, Х. и Ватанабе, Х. Дисахарид хитина (GlcNAc) 2 индуцирует естественную компетентность у Vibrio cholerae посредством транскрипционной и трансляционной активации положительного регуляторного гена tfoXVC . Gene 457 , 42–49 (2010).
CAS PubMed Google Scholar
Ямамото, С. и др. Регуляция естественной компетентности с помощью сиротской двухкомпонентной системной сенсорной киназы ChiS включает неканонический трансмембранный регулятор в Vibrio cholerae . Мол. Microbiol. 91 , 326–247 (2013).
PubMed Google Scholar
Ямамото, С.и другие. Идентификация индуцированной хитином малой РНК, которая регулирует трансляцию гена tfoX , кодирующего положительный регулятор естественной компетентности в Vibrio cholerae .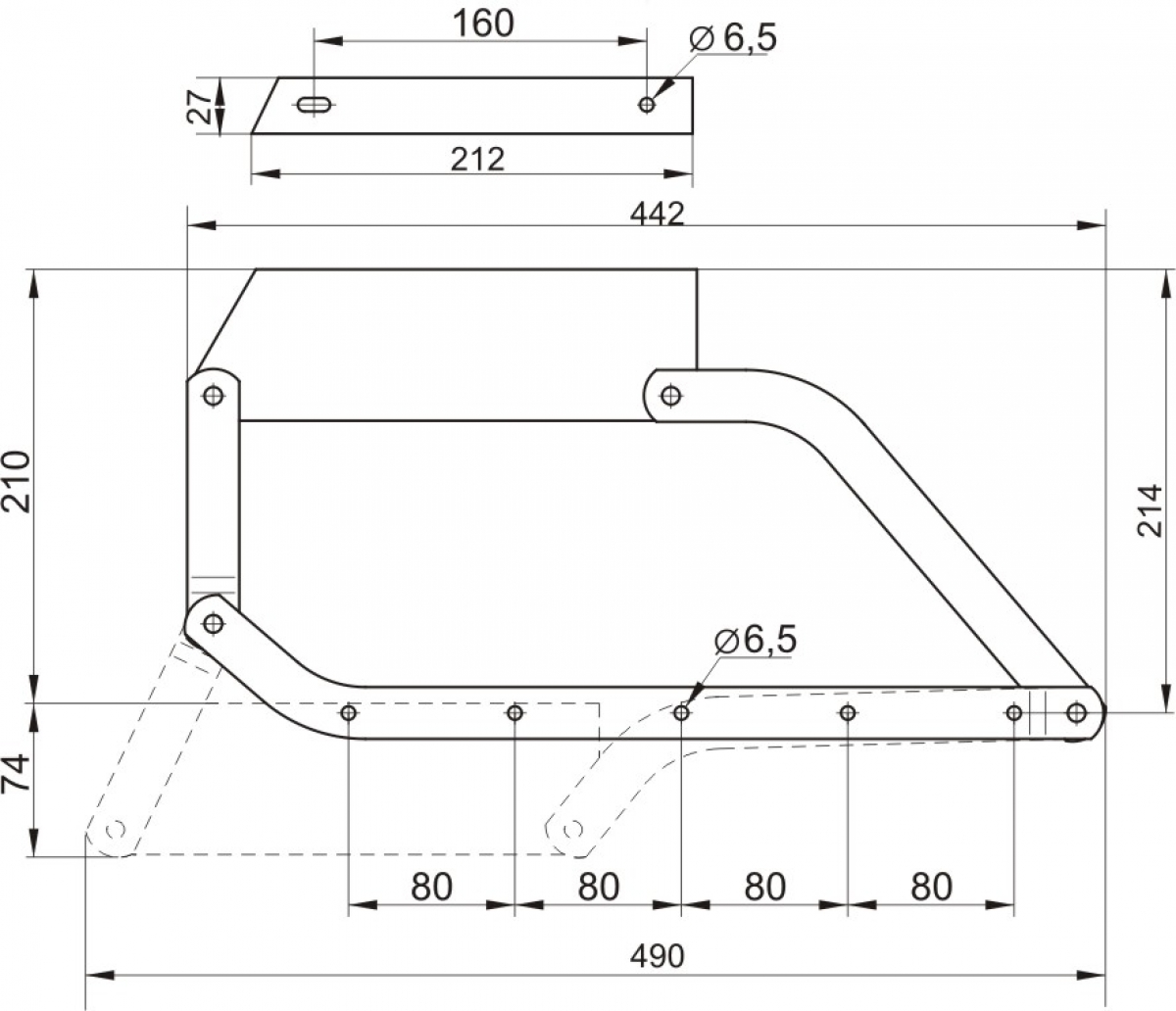 J. Bacteriol. 193 , 1953–1965 (2011).
J. Bacteriol. 193 , 1953–1965 (2011).
CAS PubMed PubMed Central Google Scholar
Херриотт, Р. М., Мейер, Э. М. и Фогт, М. Определенные среды без роста для стадии II развития компетентности у Haemophilus influenzae . J. Bacteriol. 101 , 517–524 (1970).
CAS PubMed PubMed Central Google Scholar
Bosse, J. T. et al. Естественная компетентность в отношении штаммов Actinobacillus pleuropneumoniae . FEMS Microbiol. Lett. 298 , 124–130 (2009).
CAS PubMed Google Scholar
Синха, С., Мелл, Дж.И Редфилд Р. Доступность пуриновых нуклеотидов регулирует естественную компетентность, контролируя трансляцию активатора компетентности Sxy. Мол. Microbiol. 6 , 1106–1119 (2013).
Google Scholar
Кэмерон А. Д., Волар М., Баннистер Л. А. и Редфилд Р. Дж. Вторичная структура РНК регулирует трансляцию sxy и развитие компетенции в Haemophilus influenzae . Nucleic Acids Res. 36 , 10–20 (2008).
Д., Волар М., Баннистер Л. А. и Редфилд Р. Дж. Вторичная структура РНК регулирует трансляцию sxy и развитие компетенции в Haemophilus influenzae . Nucleic Acids Res. 36 , 10–20 (2008).
CAS PubMed Google Scholar
Boutry, C. et al. Адпапторный белок MecA является негативным регулятором экспрессии генов поздней компетентности у Streptococcus thermophilus . J. Bacteriol. 194 , 1777–1788 (2012).
CAS PubMed PubMed Central Google Scholar
Биорнстад, Т.J. & Havarstein, L. S. ClpC действует как негативный регулятор компетентности в Streptococcus thermophilus . Микробиология 157 , 1676–1684 (2011).
PubMed Google Scholar
Сунг, К. и Моррисон, Д. А. Две различные функции ComW в стабилизации и активации альтернативного сигма-фактора ComX в Streptococcus pneumoniae .![]() J. Bacteriol. 187 , 3052–3061 (2005).
J. Bacteriol. 187 , 3052–3061 (2005).
CAS PubMed PubMed Central Google Scholar
Нильсен, К. М., Бонс, А. М. и ван Эльзас, Дж. Д. Вызванная естественная трансформация Acinetobacter calcoaceticus в почвенных микрокосмах. заявл. Environ. Microbiol. 63 , 3972–3977 (1997).
CAS PubMed PubMed Central Google Scholar
Ло Скрудато, М.И Блокеш М. Регуляторная сеть естественной компетентности и трансформации Vibrio cholerae . PLoS Genet. 8 , e1002778 (2012).
CAS PubMed PubMed Central Google Scholar
Кейхани Н. О. и Роземан С. Физиологические аспекты катаболизма хитина у морских бактерий. Биохим. Биофиз. Acta 1473 , 108–122 (1999).
CAS PubMed Google Scholar
Мерфи Т. Ф. и Брауэр А. Л. Экспрессия уреазы Haemophilus influenzae во время инфекции дыхательных путей человека и роль в выживании в кислой среде. BMC. Microbiol. 11 , 183 (2011).
Ф. и Брауэр А. Л. Экспрессия уреазы Haemophilus influenzae во время инфекции дыхательных путей человека и роль в выживании в кислой среде. BMC. Microbiol. 11 , 183 (2011).
CAS PubMed PubMed Central Google Scholar
Шарпантье, X., Кей, Э., Шнайдер, Д. и Шуман, Х. А. Антибиотики и УФ-излучение стимулируют способность к естественной трансформации у Legionella pneumophila . J. Bacteriol. 193 , 1114–1121 (2011).
CAS PubMed Google Scholar
Boutry, C. et al. Активация SOS-ответа и развитие компетентности являются антагонистическими механизмами у Streptococcus thermophilus . J. Bacteriol. 195 , 696–707 (2013).
CAS PubMed PubMed Central Google Scholar
Ратнаяке-Лечамвасам, М., Serror, P. , Wong, K. W. & Sonenshein, A. L. Bacillus subtilis CodY репрессирует гены ранней стационарной фазы, определяя уровни GTP. Genes Dev. 15 , 1093–1103 (2001).
, Wong, K. W. & Sonenshein, A. L. Bacillus subtilis CodY репрессирует гены ранней стационарной фазы, определяя уровни GTP. Genes Dev. 15 , 1093–1103 (2001).
CAS PubMed PubMed Central Google Scholar
Антонова, Э. С., Бернарди, Э. и Хаммер, Б. К. Естественная компетентность в отношении Vibrio cholerae контролируется реакцией поглощения нуклеозидов, которая требует CytR-зависимой антиактивации. Мол. Microbiol. 86 , 1215–1231 (2012).
CAS PubMed Google Scholar
Мичод, Р. Э., Бернштейн, Х. и Неделку, А. М. Адаптивная ценность пола в микробных патогенах. Заражение. Genet. Evol. 8 , 267–285 (2008).
CAS PubMed Google Scholar
Вос, М. Почему бактерии участвуют в гомологичной рекомбинации? Trends Microbiol.![]() 17 , 226–232 (2009).
17 , 226–232 (2009).
CAS PubMed Google Scholar
Редфилд, Р. Дж. Гены для завтрака: «съешьте и съешьте» бактериальной трансформации. J. Hered. 84 , 400–404 (1993).
CAS PubMed Google Scholar
Редфилд Р. Дж. Имеют ли бактерии половые связи? Nature Rev. Genet. 2 , 634–639 (2001).
CAS PubMed Google Scholar
Bergé, M., Mortier-Barrière, I., Martin, B. & Claverys, J.P. Трансформация Streptococcus pneumoniae основана на DprA- и RecA-зависимой защите входящих одиночных цепей. Мол. Microbiol. 50 , 527–536 (2003).
PubMed Google Scholar
Kidane, D. et al.Доказательства различных путей при горизонтальном переносе генов в компетентных клетках Bacillus subtilis . PLoS. Genet. 5 , e1000630 (2009).
PLoS. Genet. 5 , e1000630 (2009).
PubMed PubMed Central Google Scholar
Моррисон Д. А., Мортье-Баррьер И., Аттайх Л. и Клаверис Дж. П. Идентификация основного белкового компонента комплекса пневмококкового затмения. J. Bacteriol. 189 , 6497–6500 (2007).
CAS PubMed PubMed Central Google Scholar
Attaiech, L. et al. Роль одноцепочечного ДНК-связывающего белка SsbB в пневмококковой трансформации: поддержание резервуара генетической пластичности. PLoS Genet. 7 , e1002156 (2011).
CAS PubMed PubMed Central Google Scholar
Sinha, S., Mell, J.C. & Redfield, R.J. Семнадцать Sxy-зависимых генов, сайт-регулируемых белка циклического АМФ-рецептора, необходимы для естественной трансформации в Haemophilus influenzae .![]() J. Bacteriol. 194 , 5245–5254 (2012).
J. Bacteriol. 194 , 5245–5254 (2012).
CAS PubMed PubMed Central Google Scholar
Mortier-Barrière, I., de Saizieu, A., Claverys, J.P. & Martin, B. Зависящая от компетенции индукция recA требуется для полной рекомбинации во время трансформации в Streptococcus pneumoniae . Мол. Microbiol. 27 , 159–170 (1998).
PubMed Google Scholar
Lacks, SA, Ayalew, S., de la Campa, AG & Greenberg, B. Регулирование компетентности для генетической трансформации у Streptococcus pneumoniae : экспрессия dpnA , гена поздней компетентности, кодирующего ДНК-метилтрансферазу системы ограничения Dpn II. Мол. Microbiol. 35 , 1089–1098 (2000).
CAS PubMed Google Scholar
Johnston, C. et al. Запрограммированная защита чужеродной ДНК от рестрикции позволяет обменивать островки патогенности во время пневмококковой трансформации. PLoS Pathog. 9 , e1003178 (2013). Это исследование показывает, что пневмококковая метилаза, индуцированная компетентностью, имеет решающее значение для защиты трансформантных хромосом от рестрикции.
PLoS Pathog. 9 , e1003178 (2013). Это исследование показывает, что пневмококковая метилаза, индуцированная компетентностью, имеет решающее значение для защиты трансформантных хромосом от рестрикции.
CAS PubMed PubMed Central Google Scholar
Cerritelli, S., Springhorn, S. & Lacks, S.A. DpnA, метилаза для одноцепочечной ДНК в рестрикционной системе Dpn II, и ее биологическая функция. Proc. Natl Acad. Sci. США 86 , 9223–9227 (1989).
CAS PubMed Google Scholar
Джонстон, К., Мартин, Б., Полард, П. и Клаверис, Дж. П. Пострепликационное нацеливание трансформантов бактериальной иммунной системой? Trends Microbiol. 21 , 516–521 (2013).
CAS PubMed Google Scholar
Джонстон, К., Полард, П. и Клаверис, Дж. П. Пневмококковая система DpnI / DpnII, защита от внешнего нападения без нарушения генетического обмена. Моб. Genet. Элементы 3 , e25582 (2013).
П. Пневмококковая система DpnI / DpnII, защита от внешнего нападения без нарушения генетического обмена. Моб. Genet. Элементы 3 , e25582 (2013).
PubMed PubMed Central Google Scholar
Джонстон, К.и другие. Естественная генетическая трансформация создает популяцию меродиплоидов Streptococcus pneumoniae . PLoS Genet. 9 , e1003819 (2013).
CAS PubMed PubMed Central Google Scholar
Рабинович Л. и др. Иссечение профага активирует генов компетенции Listeria , которые способствуют ускользанию от фагосом и вирулентности. Cell 150 , 792–802 (2012).
CAS PubMed Google Scholar
Клэверис, Дж. П. и Мартин, Б. Гены «компетентности» бактерий: признаки активной трансформации или только остатки? Trends Microbiol.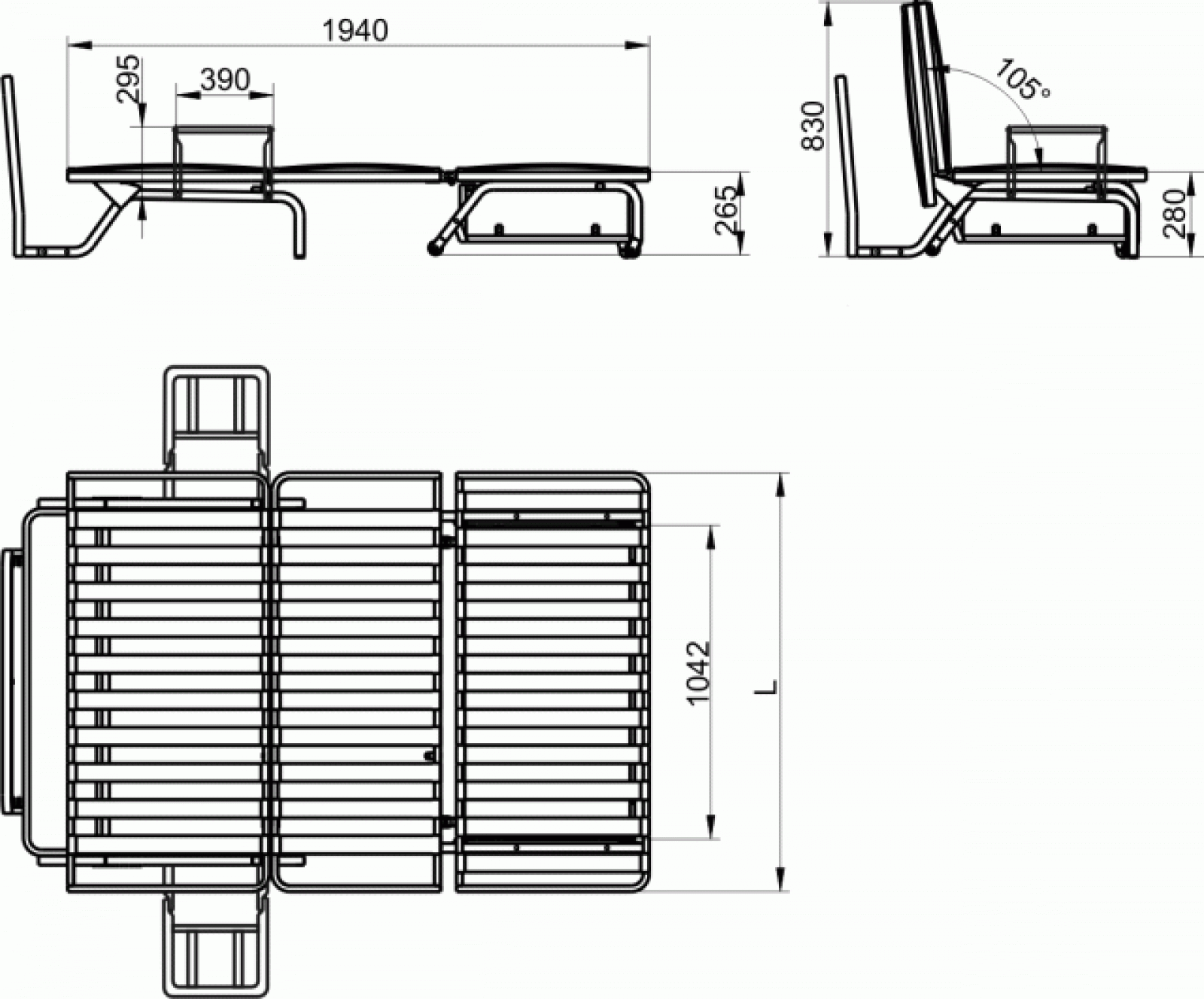 11 , 161–165 (2003).
11 , 161–165 (2003).
CAS PubMed Google Scholar
Hofreuter, D., Odenbreit, S. & Haas, R. Компетенция естественной трансформации в Helicobacter pylori опосредуется основными компонентами системы секреции типа IV. Мол. Microbiol. 41 , 379–391 (2001).
CAS PubMed Google Scholar
Karnholz, A. et al. Функциональная и топологическая характеристика новых компонентов системы компетенции comB по трансформации ДНК в Helicobacter pylori . J. Bacteriol. 188 , 882–893 (2006).
CAS PubMed PubMed Central Google Scholar
Дорер, М.С., Феро, Дж. И Салама, Н. Р. Повреждение ДНК запускает генетический обмен в Helicobacter pylori . PLoS Pathog. 6 , e1001026 (2010).
PubMed PubMed Central Google Scholar
Балтрус, Д.![]() А. и Гийемин, К. В течение цикла роста Helicobacter pylori происходит несколько фаз компетентности. FEMS Microbiol. Lett. 255 , 148–155 (2006).
А. и Гийемин, К. В течение цикла роста Helicobacter pylori происходит несколько фаз компетентности. FEMS Microbiol. Lett. 255 , 148–155 (2006).
CAS PubMed Google Scholar
Хиггинс Д.A. et al. Главный аутоиндуктор Vibrio cholerae и его роль в производстве фактора вирулентности. Природа 450 , 883–886 (2007).
CAS PubMed Google Scholar
Chen, X. et al. Структурная идентификация сигнала обнаружения кворума бактерий, содержащего бор. Nature 415 , 545–549 (2002).
CAS PubMed Google Scholar
Стивенсон, С., Мюллер, К., Цзян, М. и Перего, М. Молекулярный анализ процессинга пептида Phr в Bacillus subtilis . J. Bacteriol. 185 , 4861–4871 (2003).
CAS PubMed PubMed Central Google Scholar
Ланиган-Гердес, С.-1100x1100.jpg) , Дули, А. Н., Фаул, К. Ф. и Лазаззера, Б. А. Идентификация субтилизина, Epr и Vpr как ферментов, продуцирующих CSF, внеклеточный сигнальный пептид Bacillus subtilis . Мол. Microbiol. 65 , 1321–1333 (2007).
, Дули, А. Н., Фаул, К. Ф. и Лазаззера, Б. А. Идентификация субтилизина, Epr и Vpr как ферментов, продуцирующих CSF, внеклеточный сигнальный пептид Bacillus subtilis . Мол. Microbiol. 65 , 1321–1333 (2007).
CAS PubMed Google Scholar
Lanigan-Gerdes, S. et al. Идентификация остатков, важных для отщепления внеклеточного сигнального пептида CSF Bacillus subtilis от его белка-предшественника. J. Bacteriol. 190 , 6668–6675 (2008).
CAS PubMed PubMed Central Google Scholar
Хенке, Дж.М. и Басслер, Б. Л. Три параллельные системы контроля кворума регулируют экспрессию генов в Vibrio harveyi . J. Bacteriol. 186 , 6902–6914 (2004).
CAS PubMed PubMed Central Google Scholar
Neiditch, M. B. et al. Регулирование активности рецептора LuxPQ с помощью кворум-чувствительного сигнала аутоиндуктора-2. Мол. Ячейка 18 , 507–518 (2005).
et al. Регулирование активности рецептора LuxPQ с помощью кворум-чувствительного сигнала аутоиндуктора-2. Мол. Ячейка 18 , 507–518 (2005).
CAS PubMed Google Scholar
Ng, W.Л. и Басслер, Б. Л. Архитектуры сетей, распознающих бактериальный кворум. Annu. Преподобный Жене. 43 , 197–222 (2009).
CAS PubMed PubMed Central Google Scholar
Ленц, Д. Х. и др. Шаперон малой РНК Hfq и множественные малые РНК контролируют восприятие кворума у Vibrio harveyi и Vibrio cholerae . Cell 118 , 69–82 (2004).
CAS PubMed PubMed Central Google Scholar
Халфманн, А., Kovacs, M., Hakenbeck, R. & Bruckner, R. Идентификация генов, непосредственно контролируемых регулятором ответа CiaR в Streptococcus pneumoniae : пять из 15 промоторов управляют экспрессией малых некодирующих РНК.![]() Мол. Microbiol. 66 , 110–126 (2007).
Мол. Microbiol. 66 , 110–126 (2007).
CAS PubMed Google Scholar
Schnorpfeil, A. et al. Целевая оценка некодирующих csRNAs выявляет связь двухкомпонентной регуляторной системы CiaRH с контролем компетентности Streptococcus pneumoniae R6. Мол. Microbiol. 2 , 334–349 (2013).
Google Scholar
Стивенс, К. Э., Чанг, Д., Цвак, Э. Э. и Себерт, М. Э. Компетенция в отношении Streptococcus pneumoniae регулируется частотой ошибок декодирования рибосом. mBio 2 e00071–11 (2011).
PubMed PubMed Central Google Scholar
Кассоне, М.и другие. Протеаза HtrA из Streptococcus pneumoniae переваривает как денатурированные белки, так и пептид, стимулирующий компетентность. J. Biol. Chem. 287 , 38449–38459 (2012).
CAS PubMed PubMed Central Google Scholar
Фостер, П. Л. Стрессовые реакции и генетическая изменчивость бактерий. Mutat. Res. 569 , 3–11 (2005).
CAS PubMed PubMed Central Google Scholar
Инамин, Г.S. & Dubnau, D. ComEA, интегральный мембранный белок Bacillus subtilis , необходимый для генетической трансформации, необходим как для связывания ДНК, так и для транспорта. J. Bacteriol. 177 , 3045–3051 (1995).
CAS PubMed PubMed Central Google Scholar
Барани, Ф., Кан, М. Э. и Смит, Х. О. Направленный транспорт и интеграция донорской ДНК в Haemophilus influenzae . Proc. Natl Acad. Sci. США 80 , 7274–7278 (1983).
CAS PubMed Google Scholar
Méjean, V. & Claverys, J.P. Использование клонированного фрагмента для анализа судьбы донорской ДНК при трансформации Streptococcus pneumoniae . J. Bacteriol. 158 , 1175–1178 (1984).
& Claverys, J.P. Использование клонированного фрагмента для анализа судьбы донорской ДНК при трансформации Streptococcus pneumoniae . J. Bacteriol. 158 , 1175–1178 (1984).
PubMed PubMed Central Google Scholar
Мелл, Дж.К., Холл, И. М. и Редфилд, Р. Дж. Определение специфичности поглощения ДНК естественно компетентными клетками Haemophilus influenzae . Nucleic Acids Res. 40 , 8536–8549 (2012).
CAS PubMed PubMed Central Google Scholar
Клаверис, Дж. П. и Ховарстайн, Л. С. Каннибализм и братоубийство: механизмы и причины существования. Nature Rev. Microbiol. 5 , 219–229 (2007).
CAS Google Scholar
Клаверис, Дж. П., Мартин, Б. и Хаварштейн, Л. С. Братоубийство стрептококков, вызванное компетентностью. Мол. Microbiol. 64 , 1423–1433 (2007).
Мол. Microbiol. 64 , 1423–1433 (2007).
CAS PubMed Google Scholar
Johnsborg, O., Eldholm, V., Bjornstad, M. L. & Håvarstein, L. S. Хищный механизм резко увеличивает эффективность латерального переноса генов у Streptococcus pneumoniae и родственных комменсальных видов. Мол. Microbiol. 69 , 245–253 (2008).
CAS PubMed Google Scholar
Aas, F. E., Lovold, C. & Koomey, M. Ингибитор событий связывания и захвата ДНК определяет эффективность генетической трансформации у Neisseria gonorrhoeae : механизм действия и связь с экспрессией ворсинок IV типа. Мол. Microbiol. 46 , 1441–1450 (2002).
CAS PubMed Google Scholar
Редфилд, Р.J. et al. Эволюция компетентности и специфичности поглощения ДНК у Pasteurellaceae. BMC Evol. Биол. 6 , 82 (2006).
BMC Evol. Биол. 6 , 82 (2006).
PubMed PubMed Central Google Scholar
Фрай, С. А., Нильсен, М., Тонджум, Т. и Амбур, О. Х. Диалекты последовательности захвата ДНК в Neisseriaceae . PLoS Genet. 9 , e1003458 (2013).
CAS PubMed PubMed Central Google Scholar
Блокеш, М.И Школьник, Г. К. Внеклеточная нуклеаза Dns и ее роль в естественной трансформации Vibrio cholerae . J. Bacteriol. 190 , 7232–7240 (2008).
CAS PubMed PubMed Central Google Scholar
Финкель С. Э. и Колтер Р. ДНК как питательное вещество: новая роль гомологов генов бактериальной компетентности. J. Bacteriol. 183 , 6288–6293 (2001).
CAS PubMed PubMed Central Google Scholar
Пальчевский, В.![]() & Финкель, С.Е. Гомологи компетентных генов Escherichia coli необходимы для конкурентоспособности и использования ДНК в качестве питательного вещества. J. Bacteriol. 188 , 3902–3910 (2006).
& Финкель, С.Е. Гомологи компетентных генов Escherichia coli необходимы для конкурентоспособности и использования ДНК в качестве питательного вещества. J. Bacteriol. 188 , 3902–3910 (2006).
CAS PubMed PubMed Central Google Scholar
Сан, Д. Этюд естественной плазмидной трансформации кишечной палочки и ее взаимоотношений с программой компетентности для генетической трансформации и компетентности в питании .Диссертация, Univ. П. Сабатье, Тулуза, Франция (2011).
Google Scholar
Бломквист, Т., Стейнмоен, Х. и Ховарстайн, Л.С. Естественная генетическая трансформация: новый инструмент для эффективной генной инженерии молочной бактерии Streptococcus thermophilus . заявл. Environ. Microbiol. 72 , 6751–6756 (2006).
CAS PubMed PubMed Central Google Scholar
Морикава, К. и другие. Новый сигма-фактор стафилококка в кассете консервативных генов: функциональное значение и значение для эволюционных процессов. Genes Cells 8 , 699–712 (2003).
и другие. Новый сигма-фактор стафилококка в кассете консервативных генов: функциональное значение и значение для эволюционных процессов. Genes Cells 8 , 699–712 (2003).
CAS PubMed Google Scholar
Schmid, S., Bevilacqua, C. & Crutz-Le Coq, A. M. Альтернативный сигма-фактор σ H активирует экспрессию гена компетентности в Lactobacillus sakei . BMC.Microbiol. 12 , 32 (2012).
CAS PubMed PubMed Central Google Scholar
Опдайк, Дж. А., Скотт, Дж. Р. и Моран, К. П. Младший. Сигма-фактор вторичной РНК-полимеразы из Streptococcus pyogenes . Мол. Microbiol. 42 , 495–502 (2001).
CAS PubMed Google Scholar
Вудбери, Р.L., Wang, X. & Moran, C.P. Jr. Sigma X индуцирует экспрессию гена компетентности в Streptococcus pyogenes . Res. Microbiol. 157 , 851–856 (2006).
Res. Microbiol. 157 , 851–856 (2006).
CAS PubMed Google Scholar
Машберн-Уоррен, Л., Моррисон, Д. А. и Федерле, М. Дж. Путь криптической компетентности в Streptococcus pyogenes контролируется пептидным феромоном. J. Bacteriol. 194 , 4589–4600 (2012).
CAS PubMed PubMed Central Google Scholar
Торн К. и Стулл Х. Б. Факторы, влияющие на трансформацию Bacillus licheniformis . J. Bacteriol. 91 , 1012–1020 (1966).
CAS PubMed PubMed Central Google Scholar
Koumoutsi, A. et al. Структурная и функциональная характеристика кластеров генов, управляющих нерибосомным синтезом биоактивных циклических липопептидов в штамме Bacillus amyloliquefaciens FZB42. J. Bacteriol. 186 , 1084–1096 (2004).![]()
CAS PubMed PubMed Central Google Scholar
Ковач, А. Т., Смитс, В. К., Мирончук, А. М. и Кейперс, О. П. Повсеместно распространенные гены поздней компетентности у видов Bacillus указывают на наличие функциональных механизмов захвата ДНК. Environ. Microbiol. 11 , 1911–1922 (2009).
CAS PubMed Google Scholar
Мирончук, А.М., Ковач А. Т. и Койперс О. П. Индукция естественной компетентности у Bacillus cereus ATCC14579. Microb. Биотех. 1 , 226–235 (2008).
CAS Google Scholar
Кэмерон А. Д. и Редфилд Р. Дж. Неканонические сайты CRP контролируют регулоны компетенции в Escherichia coli и многих других γ-протеобактериях. Nucleic Acids Res. 34 , 6001–6014 (2006). Это исследование идентифицирует подмножество сайтов связывания CRP, которые зависят от регулирующего компетенцию Sxy кофактора для активации транскрипции, что привело к выводу, что sxy регулонов индуцируются во время компетенции в H.![]() influenzae , и гаммапротеобактерии.
influenzae , и гаммапротеобактерии.
CAS PubMed PubMed Central Google Scholar
Синха, С., Cameron, A.D. & Redfield, R.J. Sxy индуцирует регулон CRP-S в Escherichia coli . J. Bacteriol. 191 , 5180–5195 (2009).
CAS PubMed PubMed Central Google Scholar
Sinha, S. & Redfield, R.J. Естественное поглощение ДНК Escherichia coli . PLoS ONE. 7 , e35620 (2012).
CAS PubMed PubMed Central Google Scholar
Шимодаира, Х.И Хасегава, М. Множественные сравнения логарифма правдоподобия с приложениями к филогенетическому выводу. Мол. Биол. Evol. 16 , 1114–1116 (1999).
CAS Google Scholar
Бактериальная трансформация: механизм • Microbe Online
Бактериальная трансформация: связывание ДНК с рецепторами ДНК Последнее обновление 20 мая 2021 г.
Бактериальная трансформация — это один из трех механизмов горизонтального переноса генов, обнаруженных в бактериальных клетках.Двумя другими механизмами горизонтального переноса генов являются конъюгация (перенос плазмиды F и перенос хромосомной ДНК) и трансдукция (обобщенная трансдукция и специализированная трансдукция).
Бактериальная трансформацияОсновная информация о бактериальной трансформации
- ДНК высвобождается в окружающую среду, когда бактериальные клетки (то есть донорские) умирают и подвергаются лизису
- Реципиентные клетки поглощают свободную ДНК, выпущенную в окружающую среду.
- Не все бактерии способны извлекать свободную ДНК из окружающей среды, это могут сделать только компетентных бактерий .
Те бактерии, которые способны брать свободную ДНК и способны к трансформации, называются компетентными бактериями.
Некоторые примеры патогенных бактерий, демонстрирующих компетентность:
- Haemophilus spp
- Streptococcus spp
- Neisseria spp
Механизм трансформации 9203 в конкретном случае чтобы наблюдать, как чувствительные к ампициллину бактериальные клетки превращаются в устойчивые к ампициллину посредством поглощения гена устойчивости к ампициллину посредством трансформации.![]()
- Компетентные бактерии имеют рецепторы ДНК на поверхности клетки. Свободная ДНК, которая доступна в непосредственной близости от этих бактерий, может связываться с рецепторами ДНК.
- Двухцепочечная донорская ДНК связывается со специфическими рецепторами на поверхности компетентной клетки.
3. Только одна цепь донорной ДНК достигает внутренней части клетки-реципиента, другая цепь разрушается нуклеазой.
Бактериальная трансформация: проникновение донорской ДНК4.Происходит рекомбинация с гомологичной ДНК реципиента.
5. В игру вступает система репарации несовпадений ДНК, которая либо удаляет донорскую ДНК, либо реципиентную.
- A: Если удалить донорскую ДНК, клетка будет нести ген, чувствительный к ампициллину, но
- B: если донорская ДНК не будет удалена, клетка теперь будет иметь ген устойчивости к ампициллину.

, поэтому в смеси клеток одни клетки содержат новую донорскую ДНК, а другие — исходные последовательности ДНК.
Бактериальные клетки, которые успешно трансформируются, могут быть отобраны в лаборатории путем выращивания этих клеток в среде, содержащей ампициллин. Только клетки сопротивления будут расти.
Привет, спасибо, что посетили мой блог. Я Танкешвар Ачарья. Ведение блога — моя страсть. Я работаю ассистентом. Профессор и микробиолог отделения микробиологии и иммунологии Академии медицинских наук Патана, Непал. Если вы хотите, чтобы я писал о каких-либо постах, которые вы сочли запутанными / сложными, укажите в комментариях ниже.
7.11B: Бактериальная трансформация — Биология LibreTexts
Трансформация — это прямое поглощение, включение и экспрессия экзогенного генетического материала из окружающей среды.
ЦЕЛИ ОБУЧЕНИЯ
Различия между естественной и искусственной трансформацией
Основные выводы
Ключевые точки
- Трансформация приводит к генетическому изменению клетки-реципиента.
- Экзогенная ДНК попадает в реципиентную клетку из окружающей среды через клеточную мембрану (и).
- Трансформация происходит естественным образом у некоторых видов бактерий, но также может подвергаться искусственному воздействию в других клетках.

Ключевые термины
- эукариот : Имеющие сложные клетки, в которых генетический материал организован в мембраносвязанные ядра.
- трансформация : В молекулярной биологии трансформация — это генетическое изменение клетки, возникающее в результате прямого поглощения, включения и экспрессии экзогенного генетического материала (экзогенной ДНК) из окружающей среды и принимаемого через клеточную мембрану (мембраны).
- экспрессия : экспрессия гена — это процесс, посредством которого информация от гена используется в синтезе функционального генного продукта.
- экзогенный : Производится или происходит вне организма.
- транслоказа : фермент, который помогает перемещать другую молекулу, обычно через мембрану.

Генетическое изменение
В молекулярной биологии трансформация — это генетическое изменение клетки, возникающее в результате прямого поглощения, включения и экспрессии экзогенного генетического материала (экзогенной ДНК) из окружающей среды и принимаемого через клеточную мембрану (мембраны).
Трансформация : Иллюстрация бактериальной трансформации. ДНК мертвых клеток разрезается на фрагменты и выходит из клетки. Затем свободно плавающая ДНК может быть захвачена компетентными клетками. Экзогенная ДНК включается в хромосому клетки-хозяина посредством рекомбинации.
ПРИРОДНОЕ ПРЕОБРАЗОВАНИЕ
Трансформация происходит естественным образом у некоторых видов бактерий, но также может быть осуществлена искусственным путем в других клетках. Чтобы трансформация произошла, бактерии должны быть в состоянии компетентности, что может происходить как ограниченная по времени реакция на условия окружающей среды, такие как голодание и плотность клеток.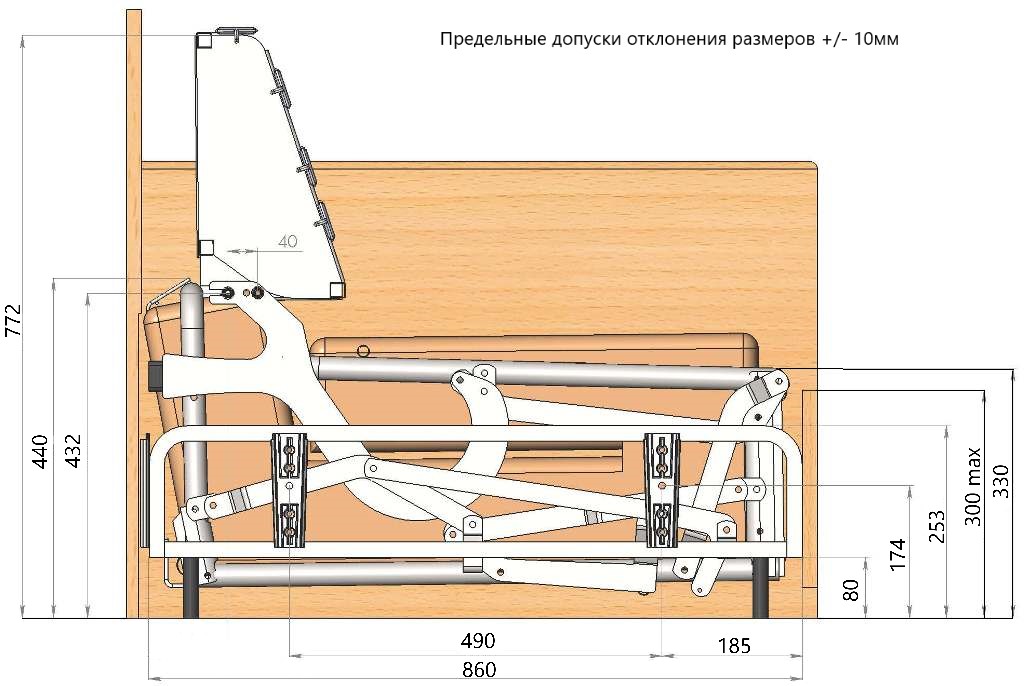 Трансформация — это один из трех процессов, посредством которых экзогенный генетический материал может быть введен в бактериальную клетку; два других — конъюгация (перенос генетического материала между двумя бактериальными клетками в прямом контакте) и трансдукция (инъекция чужеродной ДНК вирусом бактериофага в бактерию-хозяин).
Трансформация — это один из трех процессов, посредством которых экзогенный генетический материал может быть введен в бактериальную клетку; два других — конъюгация (перенос генетического материала между двумя бактериальными клетками в прямом контакте) и трансдукция (инъекция чужеродной ДНК вирусом бактериофага в бактерию-хозяин).
« Трансформация » также может использоваться для описания внедрения нового генетического материала в небактериальные клетки, включая клетки животных и растений; однако, поскольку «трансформация» имеет особое значение по отношению к животным клеткам, указывая на прогрессирование до ракового состояния, следует избегать этого термина для животных клеток при описании введения экзогенного генетического материала.Введение чужеродной ДНК в эукариотические клетки часто называют «трансфекцией ».
Бактериальную трансформацию можно назвать стабильным генетическим изменением, вызванным поглощением «голой» ДНК (ДНК без ассоциированных клеток или белков). Компетентность относится к состоянию способности поглощать экзогенную ДНК из окружающей среды. Есть две формы компетенции: естественная и искусственная.
Компетентность относится к состоянию способности поглощать экзогенную ДНК из окружающей среды. Есть две формы компетенции: естественная и искусственная.
Около 1% видов бактерий способны естественным образом поглощать ДНК в лабораторных условиях; большее количество людей может принять это в своей естественной среде обитания.Материал ДНК может передаваться между различными штаммами бактерий в процессе, который называется горизонтальным переносом генов.
Некоторые виды после гибели клетки высвобождают свою ДНК, которая будет поглощена другими клетками; однако трансформация лучше всего работает с ДНК близкородственных видов. Эти естественно компетентные бактерии несут наборы генов, которые обеспечивают белковый аппарат для переноса ДНК через клеточную мембрану (и). Для транспорта экзогенной ДНК в клетки могут потребоваться белки, которые участвуют в сборке пилей типа IV и системы секреции типа II, а также комплекса транслоказ ДНК на цитоплазматической мембране.
РАЗЛИЧИЯ ПОЛОЖИТЕЛЬНОЙ И ОТРИЦАТЕЛЬНОЙ ПО ГРАММАМ
Из-за различий в структуре клеточной оболочки грамположительных и грамотрицательных бактерий существуют некоторые различия в механизмах захвата ДНК этими клетками. Однако большинство из них имеют общие черты, связанные с родственными белками. ДНК сначала связывается с поверхностью компетентных клеток на рецепторе ДНК и проходит через цитоплазматическую мембрану через ДНК-транслоказу. Может проходить только одноцепочечная ДНК, поэтому одна цепь разрушается нуклеазами в этом процессе, и транслоцированная одноцепочечная ДНК может затем интегрироваться в бактериальные хромосомы посредством RecA-зависимого процесса.
Однако большинство из них имеют общие черты, связанные с родственными белками. ДНК сначала связывается с поверхностью компетентных клеток на рецепторе ДНК и проходит через цитоплазматическую мембрану через ДНК-транслоказу. Может проходить только одноцепочечная ДНК, поэтому одна цепь разрушается нуклеазами в этом процессе, и транслоцированная одноцепочечная ДНК может затем интегрироваться в бактериальные хромосомы посредством RecA-зависимого процесса.
В грамотрицательных клетках из-за наличия дополнительной мембраны ДНК требует наличия канала, образованного секретинами на внешней мембране. Пилин может потребоваться для компетентности, однако его роль неясна. Поглощение ДНК обычно неспецифично для последовательности, хотя у некоторых видов присутствие конкретных последовательностей захвата ДНК может способствовать эффективному захвату ДНК.
ИСКУССТВЕННАЯ ПЕРЕДАЧА
Искусственная компетентность может быть вызвана лабораторными процедурами, которые включают создание пассивной проницаемости клетки для ДНК путем воздействия на нее условий, которые обычно не встречаются в природе.![]() Обычно клетки инкубируют в растворе, содержащем двухвалентные катионы; чаще всего раствор хлорида кальция в холодных условиях, который затем подвергается тепловому шоку. Однако механизм захвата ДНК посредством химически индуцированной компетентности в этом методе трансформации хлорида кальция неясен.
Обычно клетки инкубируют в растворе, содержащем двухвалентные катионы; чаще всего раствор хлорида кальция в холодных условиях, который затем подвергается тепловому шоку. Однако механизм захвата ДНК посредством химически индуцированной компетентности в этом методе трансформации хлорида кальция неясен.
Поверхность бактерий, таких как E. coli, заряжена отрицательно из-за фосфолипидов и липополисахаридов на ее клеточной поверхности, а ДНК также заряжена отрицательно.Таким образом, одной из функций двухвалентного катиона является экранирование зарядов путем координации фосфатных групп и других отрицательных зарядов, тем самым позволяя молекуле ДНК прикрепляться к поверхности клетки. Предполагается, что воздействие на клетки двухвалентных катионов в холодных условиях может также изменить или ослабить структуру клеточной поверхности клеток, делая ее более проницаемой для ДНК. Считается, что тепловой импульс создает тепловой дисбаланс с обеих сторон клеточной мембраны, что заставляет ДНК проникать в клетки либо через клеточные поры, либо через поврежденную клеточную стенку.
Электропорация — еще один метод повышения компетентности. Используя этот метод, клетки кратковременно подвергают воздействию электрического поля 10-20 кВ / см, которое, как считается, создает отверстия в клеточной мембране, через которые может проникнуть плазмидная ДНК. После поражения электрическим током отверстия быстро закрываются механизмами восстановления мембраны клетки.
O. T. Avery, et al. были первыми, кто продемонстрировал, что «грубые» колонии S. pneumoniae могут быть преобразованы в «гладкие» (образующие капсулы) колонии путем добавления экстрактов ДНК первых ко вторым, таким образом «трансформируя» их.(См. Ледерберг ниже)
- Ледерберг, Джошуа (1994). Трансформация генетики с помощью ДНК: празднование годовщины AVERY, MACLEOD и MCCARTY (1944) в анекдотических, исторических и критических комментариях по генетике. Университет Рокфеллера, Нью-Йорк, Нью-Йорк 10021-6399. PMID 8150273.
границ | Пересмотр механизмов, вовлеченных в бактериальную трансформацию, индуцированную хлоридом кальция
Бактериальная трансформация является важной частью процесса клонирования и широко используется во многих исследованиях (Swords, 2003; Gigova et al.![]() , 2006). Этот механизм отмечен двумя фазами: первая фаза включает поглощение ДНК через клеточную оболочку, а вторая фаза включает создание ДНК в клетке как стабильного генетического материала (Hanahan, 1983). Процедура трансформации носит физико-химический характер, а не является строго химической или физической процедурой, поскольку клетки обрабатываются катионами (или комбинацией катионов) и температурным дисбалансом, чтобы сделать их способными поглощать чужеродную ДНК.Принято считать, что химические манипуляции связаны с индукцией компетентности, в то время как физические манипуляции связаны с поглощением чужеродной ДНК. Эти два метода действуют вместе, чтобы вызвать акт «искусственной интернализации ДНК».
, 2006). Этот механизм отмечен двумя фазами: первая фаза включает поглощение ДНК через клеточную оболочку, а вторая фаза включает создание ДНК в клетке как стабильного генетического материала (Hanahan, 1983). Процедура трансформации носит физико-химический характер, а не является строго химической или физической процедурой, поскольку клетки обрабатываются катионами (или комбинацией катионов) и температурным дисбалансом, чтобы сделать их способными поглощать чужеродную ДНК.Принято считать, что химические манипуляции связаны с индукцией компетентности, в то время как физические манипуляции связаны с поглощением чужеродной ДНК. Эти два метода действуют вместе, чтобы вызвать акт «искусственной интернализации ДНК».
Со временем для придания клеткам компетентности использовались различные методы, такие как использование диметилсульфоксида (ДМСО), двухвалентных катионов или полиэтиленгликоля (ПЭГ) (Klebe et al., 1983; Chan et al., 2013). Помимо этих химических методов, электропорация также была протестирована и использовалась для повышения компетентности (Dower et al. , 1988; Ёсида и Сато, 2009 г .; Лю и др., 2013). Использование двухвалентных катионов было наиболее эффективной химической обработкой для трансформации (Day, 2004). Среди различных катионов двухвалентный катион кальция (Ca 2+ ) оказался наиболее эффективным (Weston et al., 1981) как сам по себе (Dagert and Ehrlich, 1979), так и в различных комбинациях. Комбинация двухвалентных и одновалентных ионов, таких как кальций и магний (Taketo, 1974; Wensink et al., 1974), кальций и марганец (Enea et al., 1975), кальций, рубидий и диметилсульфоксид (Kushner, 1978) и другие щелочные металлы при длительной инкубации при 0 ° C (Taketo, 1972; Dagert and Ehrlich, 1979) также считаются эффективными (Roychoudhury et al. ., 2009). Как правило, все двухвалентные катионы усиливают процесс превращения. Ханахан (1983) обнаружил, что присутствие магния в бактериальной культуральной среде увеличивает эффективность трансформации в 15-20 раз по сравнению с клетками, выращенными в отсутствие магния. Он также заметил, что добавление магния в среду за 30 мин до момента сбора клеток также увеличивает трансформацию до ~ 60%.Однако добавление магния во время сбора и инкубации клеток на льду увеличивает трансформацию до ~ 40%. Добавление ионов кальция или марганца также показало почти такой же стимулирующий эффект, что и ионы магния (Hanahan, 1983). Однако следует оптимизировать время инкубации с хлоридом кальция или любым другим двухвалентным катионом. Было замечено, что период инкубации в течение 24 часов в холодном хлориде кальция делает бактериальные клетки в 20-30 раз более компетентными и в 4-6 раз более эффективными для трансформации по сравнению с клетками, полученными сразу после обработки CaCl 2 (Blattner и другие., 1977; Дагерт и Эрлих, 1979). Curtiss et al. (1977) проводили эксперименты на штамме X1776 E. coli и наблюдали влияние различных условий на эффективность трансформации. Он обработал бактериальную культуру комбинацией ионов марганца, кальция, рубидия и калия вместе с ДМСО и сахарозой при 0 ° C с последующим тепловым импульсом при 42 ° C. Однако эти условия не дали успешных результатов при использовании других штаммов E. coli (Hanahan, 1983). Мезельсон и Юань (1968) сочли эти условия многообещающими для успешной трансформации штамма E.coli MM294, чем стандартный протокол «хлорид кальция». Bolivar et al. (1977) сообщили о 10 6 трансформантах, когда клетки обрабатывали одним кальцием, в то время как Кушнер (1978) сообщил о получении 10 7 трансформантов после обработки клеток рубидием вместе с хлоридом кальция. Norgard et al. (1978) также удалось получить 10 7 трансформантов с помощью метода, использованного Кушнером (1978) в случае штамма K-12 X1776 из E. coli . Однако выход трансформантов варьировал от вида к виду и от штамма к штамму (Mercer and Loutit, 1979).Об этом сообщили Sjöström et al. (1972), что оптимальная концентрация хлорида кальция для поглощения ДНК S. aureus составляет 0,1 М CaCl 2 . Эти двухвалентные катионы преодолевают отталкивание между чужеродной ДНК и бактериальной клеткой из-за отрицательного заряда на них обоих. Это применимо как для линейных фрагментов ДНК, так и для кольцевых молекул ДНК, таких как плазмиды (Mandel and Higa, 1970; Tsen et al., 2002). Считается, что двухвалентные катионы связываются как с клеткой, так и с ДНК, тем самым полностью нейтрализуя заряд.Связанный с ДНК кальций дополнительно помогает ДНК адсорбироваться в компетентной клетке (Panja et al., 2008b). Более того, ДНК-связывающие белки, присутствующие в клеточной мембране, также могут способствовать этому взаимодействию. Закрепление ДНК на мембране устраняет риск отделения или изгнания ДНК (Clark et al., 2002). Кроме того, низкие температуры, используемые в протоколах трансформации, застывают липидный фрагмент и, следовательно, ограничивают текучесть клеточной мембраны, что усиливает взаимодействие кальция с поверхностью клетки.Таким образом, ионы кальция, связанные с поверхностью клетки, а также с чужеродной ДНК, переносят ДНК в клетку. Кларк и др. (2002) показали, что относительная ассоциация двухвалентных катионов (например, Ca 2+ ) больше связана с клеточной мембраной по сравнению с его ассоциацией с чужеродной ДНК, тогда как некоторые трехвалентные катионы (например, спермидин) легче взаимодействуют с ДНК. (Ли и др., 2004). В этом исследовании также сообщалось, что Ca 2+ играет более выраженную роль в развитии компетентности по сравнению со спермидином или трехвалентными катионами (Clark et al., 2002). Мембраны очень легко поглощают кальций, и, оказавшись внутри клетки, ионы кальция нейтрализуются мембранными фосфатами, присутствующими на цитозольной стороне (Melcrová et al., 2016). Связывание ионов кальция с мембраной также вызывает изменения проницаемости мембраны (Li et al., 2004).
, 1988; Ёсида и Сато, 2009 г .; Лю и др., 2013). Использование двухвалентных катионов было наиболее эффективной химической обработкой для трансформации (Day, 2004). Среди различных катионов двухвалентный катион кальция (Ca 2+ ) оказался наиболее эффективным (Weston et al., 1981) как сам по себе (Dagert and Ehrlich, 1979), так и в различных комбинациях. Комбинация двухвалентных и одновалентных ионов, таких как кальций и магний (Taketo, 1974; Wensink et al., 1974), кальций и марганец (Enea et al., 1975), кальций, рубидий и диметилсульфоксид (Kushner, 1978) и другие щелочные металлы при длительной инкубации при 0 ° C (Taketo, 1972; Dagert and Ehrlich, 1979) также считаются эффективными (Roychoudhury et al. ., 2009). Как правило, все двухвалентные катионы усиливают процесс превращения. Ханахан (1983) обнаружил, что присутствие магния в бактериальной культуральной среде увеличивает эффективность трансформации в 15-20 раз по сравнению с клетками, выращенными в отсутствие магния. Он также заметил, что добавление магния в среду за 30 мин до момента сбора клеток также увеличивает трансформацию до ~ 60%.Однако добавление магния во время сбора и инкубации клеток на льду увеличивает трансформацию до ~ 40%. Добавление ионов кальция или марганца также показало почти такой же стимулирующий эффект, что и ионы магния (Hanahan, 1983). Однако следует оптимизировать время инкубации с хлоридом кальция или любым другим двухвалентным катионом. Было замечено, что период инкубации в течение 24 часов в холодном хлориде кальция делает бактериальные клетки в 20-30 раз более компетентными и в 4-6 раз более эффективными для трансформации по сравнению с клетками, полученными сразу после обработки CaCl 2 (Blattner и другие., 1977; Дагерт и Эрлих, 1979). Curtiss et al. (1977) проводили эксперименты на штамме X1776 E. coli и наблюдали влияние различных условий на эффективность трансформации. Он обработал бактериальную культуру комбинацией ионов марганца, кальция, рубидия и калия вместе с ДМСО и сахарозой при 0 ° C с последующим тепловым импульсом при 42 ° C. Однако эти условия не дали успешных результатов при использовании других штаммов E. coli (Hanahan, 1983). Мезельсон и Юань (1968) сочли эти условия многообещающими для успешной трансформации штамма E.coli MM294, чем стандартный протокол «хлорид кальция». Bolivar et al. (1977) сообщили о 10 6 трансформантах, когда клетки обрабатывали одним кальцием, в то время как Кушнер (1978) сообщил о получении 10 7 трансформантов после обработки клеток рубидием вместе с хлоридом кальция. Norgard et al. (1978) также удалось получить 10 7 трансформантов с помощью метода, использованного Кушнером (1978) в случае штамма K-12 X1776 из E. coli . Однако выход трансформантов варьировал от вида к виду и от штамма к штамму (Mercer and Loutit, 1979).Об этом сообщили Sjöström et al. (1972), что оптимальная концентрация хлорида кальция для поглощения ДНК S. aureus составляет 0,1 М CaCl 2 . Эти двухвалентные катионы преодолевают отталкивание между чужеродной ДНК и бактериальной клеткой из-за отрицательного заряда на них обоих. Это применимо как для линейных фрагментов ДНК, так и для кольцевых молекул ДНК, таких как плазмиды (Mandel and Higa, 1970; Tsen et al., 2002). Считается, что двухвалентные катионы связываются как с клеткой, так и с ДНК, тем самым полностью нейтрализуя заряд.Связанный с ДНК кальций дополнительно помогает ДНК адсорбироваться в компетентной клетке (Panja et al., 2008b). Более того, ДНК-связывающие белки, присутствующие в клеточной мембране, также могут способствовать этому взаимодействию. Закрепление ДНК на мембране устраняет риск отделения или изгнания ДНК (Clark et al., 2002). Кроме того, низкие температуры, используемые в протоколах трансформации, застывают липидный фрагмент и, следовательно, ограничивают текучесть клеточной мембраны, что усиливает взаимодействие кальция с поверхностью клетки.Таким образом, ионы кальция, связанные с поверхностью клетки, а также с чужеродной ДНК, переносят ДНК в клетку. Кларк и др. (2002) показали, что относительная ассоциация двухвалентных катионов (например, Ca 2+ ) больше связана с клеточной мембраной по сравнению с его ассоциацией с чужеродной ДНК, тогда как некоторые трехвалентные катионы (например, спермидин) легче взаимодействуют с ДНК. (Ли и др., 2004). В этом исследовании также сообщалось, что Ca 2+ играет более выраженную роль в развитии компетентности по сравнению со спермидином или трехвалентными катионами (Clark et al., 2002). Мембраны очень легко поглощают кальций, и, оказавшись внутри клетки, ионы кальция нейтрализуются мембранными фосфатами, присутствующими на цитозольной стороне (Melcrová et al., 2016). Связывание ионов кальция с мембраной также вызывает изменения проницаемости мембраны (Li et al., 2004).
За обработкой дивалентами или тривалентами на льду следует обработка при повышенной температуре в качестве теплового шока, вызывающего температурный дисбаланс. Молекулы с усиленным броуновским движением вне клетки, вероятно, вытеснят молекулу ДНК внутрь клетки.Однако неясно, достаточно ли этой кинетической силы, чтобы протолкнуть адсорбированные молекулы ДНК внутрь. Panja et al. (2008a) изучали эффективность циклов охлаждения и нагрева, увеличивая количество циклов до достижения максимальной эффективности преобразования. Был сделан вывод, что снижение температуры действительно способствует потере белка, тогда как нагревание способствует потере липидов, и, таким образом, вместе эти циклы увеличивают эффективность трансформации (Panja et al., 2008a), поскольку это увеличивает размер пор на поверхности клетки.Более того, из-за потери липидов и белков мембрана деполяризуется, что еще больше снижает отталкивание между молекулой ДНК и мембраной. Более того, плотность клеток также может влиять на эффективность трансформации, и сообщалось, что максимальная компетентность наблюдается при плотности клеток в диапазоне от 10 7 до 10 8 клеток на мл в логарифмической фазе (Taketo, 1974; Norgard et al. , 1978).
Однако остается вопрос; сформированы ли поры (через которые чужеродная ДНК проникает в клетку) обработкой кальцием или они присутствуют в природе.В мембране существуют естественные каналы, часто называемые мостиками Байера, которые могут служить потенциальным путем для захвата ДНК (Dreiseikelmann, 1994; Sperandeo et al., 2007; Srivastava, 2013). Ханахан (1983) заявил, что компетентные клетки имеют много участков или каналов, и все эти участки и каналы имеют независимый шанс принять участие в поглощении ДНК, продвигаясь к процессу трансформации. Все клетки, независимо от того, являются они компетентными или нет, конкурируют за захват плазмиды, но если для трансформации используются только компетентные клетки, эффективность будет увеличена до 50 раз, как обсуждалось Hanahan (1983).Фактор поглощения ДНК — это сумма всех вероятностей поглощения ДНК через каждый канал. Сообщалось, что шансы на трансформацию увеличиваются не за счет увеличения концентрации ДНК, а за счет увеличения количества каналов, через которые происходит поглощение ДНК (Hanahan, 1983; Nikaido and Vaara, 1985). Более того, кальций играет в этом процессе двоякую роль; он не только нейтрализует заряд, но также ослабляет клеточную мембрану, вызывая инвагинации (Stein, 1990; Thomas and Rice, 2014).
Хотя было известно, что двухвалентные катионы помогают нейтрализовать заряд, сложные ионы также могут служить для создания статической силы притяжения внутри молекулы ДНК. Это приводит к сворачиванию ДНК в компактную шарообразную структуру, которая облегчает ее проникновение в клетку (Clark et al., 2002). Сверхспиральная шарообразная структура плазмиды будет иметь больше шансов попасть в компетентную клетку для трансформации, чем расширенная открытая круглая форма плазмиды. Однако, если размер ДНК приближается к размеру поры, вероятность трансформации резко снижается.При использовании спермидина или других тривалентов размер шарообразной структуры ДНК может превышать размер пор в клеточной мембране, что может быть решено только путем изменения физических параметров, используемых в протоколе, в первую очередь циклов нагрева и охлаждения. Независимо от того, используются ли диваленты или триваленты, их концентрации должны быть оптимизированы так, чтобы все фосфаты ДНК не становились недоступными, потому что некоторые части ДНК должны адсорбироваться на поверхности клетки, а для этого требуются свободные фосфаты, как считает Панджа. и другие.(2008b).
На эффективность трансформации сильно влияет тип клетки-хозяина, поскольку они имеют разные структуры клеточной поверхности, особенно в отношении О-полисахаридов, которые выступают с поверхности клетки. Эти поверхностные структуры взаимодействуют с двухвалентными катионами и ДНК, что делает клетку пригодной для трансформации. Сообщалось, что различные штаммы E. coli , как обсуждалось выше, демонстрируют различия в эффективности трансформации из-за различий в химических свойствах их клеточной оболочки (Taketo, 1972).Очень плотный О-полисахарид станет препятствием для прохождения ДНК. Однако также утверждалось, что обширное удаление LPS путем чрезмерной предварительной обработки этанолом снижает эффективность трансформации (Roychoudhury et al., 2009). Это можно объяснить вышеупомянутой гипотезой о том, что ДНК сначала прикрепляется к некоторому внешнему компоненту клеточной мембраны, который затем помогает ей перемещаться внутри клетки. Наряду с плотностью состав О-полисахарида также играет роль в рецепции входящей молекулы ДНК (Lacks, 1977).Более того, ионы кальция также взаимодействуют с мембраной, и при концентрации 100 мМ CaCl 2 почти весь кальций поглощается фосфатидилхолином и фосфатидилсерином клеточной мембраны (Melcrová et al., 2016). Следовательно, свойства мембраны играют важную роль в адсорбции ДНК.
Доказательства ясно показывают, что физические и химические методы обработки, используемые во время трансформации, то есть температурный дисбаланс и обработка CaCl 2 , помогают справиться с барьерами для поглощения ДНК, такими как отталкивание зарядов и размер пор (рис.Комбинации магния и кальция редко используются в протоколах трансформации, важность которых следует учитывать. Комбинация двухвалентных и трехвалентных катионов с увеличенным временем инкубации может быть предложена для повышения эффективности трансформации; поскольку в дополнение к стабилизации заряда трехвалентные катионы могут уплотнять ДНК, способствуя ее интернализации. Бактериальные клетки также можно было выращивать в присутствии CaCl 2 и MgCl 2 перед тем, как вызвать компетентность.Циклы нагрева и охлаждения, используемые только один раз в протоколах преобразования, также могут быть увеличены до трех раз для повышения эффективности преобразования. Эти условия необходимо отрегулировать и оптимизировать для различных видов и штаммов бактерий из-за различий в их поверхностных свойствах. Однако необходимы конкретные доказательства, основанные на экспериментах, разработанных исключительно для изучения этого явления.
Рисунок 1 . показывает барьеры / ограничения в поглощении ДНК бактериальной клеткой, которые есть; отталкивание, вызванное отрицательными зарядами на клеточной мембране и ДНК, а также пористость мембраны.Они обрабатываются химической обработкой, такой как ионы кальция, которые нейтрализуют отрицательные заряды. Физические параметры могут применяться для улучшения пористости и проницаемости.
Авторские взносы
AA и HM составили рукопись. YR выдвинул идею рукописи и отредактировал рукопись до окончательной формы. RT помог в написании рукописи.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Список литературы
Блаттнер, Ф. Р., Уильямс, Б. Г., Блехл, А. Э., Деннистон-Томпсон, К., Фабер, Х. Э., Ферлонг, Л. и др. (1977). Фаги Харона: более безопасные производные бактериофага лямбда для клонирования ДНК. Наука 196, 161–169. DOI: 10.1126 / science.847462
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Боливар, Ф., Родригес, Р. Л., Грин, П. Дж., Бетлах, М. К., Хейнекер, Х. Л., Бойер, Х. В. и др. (1977). Построение и описание новой машины для клонирования.II. Многоцелевая система клонирования. Gene 2, 95–113.
PubMed Аннотация
Чан, В.-Т., Верма, С.С., Лейн, Д.П., и Ган, С.К. (2013). Сравнение и оптимизация методов и факторов, влияющих на трансформацию Escherichia coli . Biosci. Реп. 33: e00086. DOI: 10.1042 / BSR20130098
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кларк, Дж., Хадсон, Дж., Мак, Р., Макферсон, К., и Цин, К. (2002).Анализ эффективности трансформации в E. coli : исследование относительной эффективности E. coli по захвату плазмидной ДНК, обработанной сложными молекулярными трехвалентными катионами спермина или спермидина в контексте протокола Hanahan для трансформации. J. Exp. Microbiol. Иммунол . 2, 68–80.
Google Scholar
Curtiss, R. III, Inoue, M., Pereira, D., Hsu, J. C., Alexander, L., and Rock, L. (1977). «Создание и использование более безопасных бактериальных штаммов-хозяев для исследования рекомбинантной ДНК», в Molecular Cloning of Recombinant DNA , eds W.А. Скотт и Р. Вернер (Нью-Йорк, Нью-Йорк: Academic Press), 99–114.
Google Scholar
Дэй, М. Дж. (2004). «Трансформация», в Microbial Evolution , ред. Р. В. Миллер и М. Дж. Дэй (Вашингтон, округ Колумбия: Издательство Американского общества микробиологии), 158–172.
Google Scholar
Дауэр, У. Дж., Миллер, Дж. Ф. и Рэгсдейл, К. У. (1988). Высокоэффективная трансформация E. coli с помощью электропорации под высоким напряжением. Nucleic Acids Res .16, 6127–6145. DOI: 10.1093 / nar / 16.13.6127
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Энеа В., Вовис Г. П. и Зиндер Н. Д. (1975). Генетические исследования с гетеродуплексной ДНК бактериофага f1. Асимметричная сегрегация, коррекция оснований и значение механизма генетической рекомбинации. J. Mol. Биол. 96, 495–509. DOI: 10.1016 / 0022-2836 (75) -8
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гигова, Л., Петрова, Н., Димова, С., Станева, Д. (2006). Клонирование nifM-подобного гена из Sinorhizobium meliloti . Comptes rendus de l’Acad’emie bulgare des Sci. 59, 865–868.
Google Scholar
Клебе, Р. Дж., Харрисс, Дж. В., Шарп, З. Д., и Дуглас, М. Г. (1983). Общий метод генетической трансформации бактерий и дрожжей, индуцированной полиэтиленгликолем. Gene 25, 333–341. DOI: 10.1016 / 0378-1119 (83)
-XPubMed Аннотация | CrossRef Полный текст | Google Scholar
Кушнер, С.Р. (1978). Улучшенный метод трансформации Escherichia coli плазмидами, производными ColEl . Амстердам: Эльзевир.
Лакс, С. А. (1977). Связывание и участие ДНК в микробных взаимодействиях трансформации бактерий (Рецепторы и распознавание, серия, Б., том 3). Лондон: Чепмен и Холл.
Ли, В., Се, Х., Се, З., Лу, З., Оу, Дж., Чен, X., и др. (2004). Изучение механизма развития компетентности у Escherichia coli с использованием квантовых точек в качестве флуоресцентных зондов. J. Biochem. Биофиз. Методы 58, 59–66. DOI: 10.1016 / S0165-022X (03) 00154-4
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лю К., Се X., Чжао В., Лю Н., Маракчини П. А., Сассубр Л. М. и др. (2013). Проведение электропорации с наношубками для доступной и высокоэффективной дезинфекции воды от бактерий и вирусов. Nano Lett. 13, 4288–4293. DOI: 10.1021 / nl402053z
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мелькрова, А., Pokorna, S., Pullanchery, S., Kohagen, M., Jurkiewicz, P., Hof, M., et al. (2016). Сложный характер взаимодействия катиона кальция с бислоями фосфолипидов. Sci. Реп. 6: 38035. DOI: 10.1038 / srep38035
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мерсер А. А. и Лутит Дж. С. (1979). Трансформация и трансфекция Pseudomonas aeruginosa : эффекты ионов металлов. J. Bacteriol. 140, 37–42.
PubMed Аннотация | Google Scholar
Никайдо, Х., и Ваара, М. (1985). Молекулярные основы проницаемости внешней мембраны бактерий. Microbiol. Ред. 49, 1.
PubMed Аннотация | Google Scholar
Норгард, М. В., Ким, К., и Монахан, Дж. Дж. (1978). Факторы, влияющие на трансформацию штамма Escherichia coli χ1776 плазмидной ДНК pBR322. Gene 3, 279–292. DOI: 10.1016 / 0378-1119 (78)
-0PubMed Аннотация | CrossRef Полный текст | Google Scholar
Panja, S., Aich, P., Яна Б. и Басу Т. (2008a). Как плазмидная ДНК проникает через клеточные мембраны в процессе искусственной трансформации Escherichia coli ? Мол. Membr. Биол. 25, 411–422. DOI: 10.1080 / 09687680802187765
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Панджа, С., Айч, П., Яна, Б., и Басу, Т. (2008b). Плазмидная ДНК связывается с коровым олигосахаридным доменом молекул LPS клеточной поверхности E. coli в процессе трансформации, опосредованной CaCl2. Биомакромолекулы 9, 2501–2509. DOI: 10.1021 / bm8005215
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ройчоудхури А., Басу С. и Сенгупта Д. Н. (2009). Анализ сравнительной эффективности различных методов трансформации E. coli с использованием двух общих плазмидных векторов. Indian J. Biochem. Биофиз . 46, 395–400.
PubMed Аннотация | Google Scholar
Sjöström, J.-E., Lindberg, M., and Philipson, L.(1972). Трансфекция Staphylococcus aureus бактериофагом дезоксирибонуклеиновой кислотой. J. Bacteriol. 109, 285–291.
PubMed Аннотация | Google Scholar
Сперандео, П., Ческутти, Р., Вилья, Р., Ди Бенедетто, К., Кандиа, Д., Дехо, Г. и др. (2007). Характеристика lptA и lptB, двух основных генов, участвующих в транспорте липополисахаридов к внешней мембране Escherichia coli . J. Bacteriol. 189, 244–253. DOI: 10.1128 / JB.01126-06
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Шривастава, С. (2013). Преобразование. Генетика бактерий. Нью-Дели: Springer India.
Штейн, С. (1990). «Производство и анализ белков с помощью технологии рекомбинантных ДНК», в Основы белковой биотехнологии (CRC Press).
Google Scholar
Мечи, У. Э. (2003). «Химическая трансформация E. coli », в E.coli Plasmid Vectors: Methods and Applications , ред. Н. Касали и А. Престон (Тотова, Нью-Джерси; Оксфорд: Humana; Blackwell), 49–53.
Google Scholar
Такето, А. (1972). Чувствительность Escherichia coli к вирусной нуклеиновой кислоте: v. Компетентность клеток, обработанных кальцием. Дж. Биохим . 72, 973–979. DOI: 10.1093 / oxfordjournals.jbchem.a129988
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Цен, С.-Д., Фанг, С.-С., Чен, М.-J., Chien, J.-Y., Lee, C.-C., и Tsen, D.H. (2002). Трансформация естественной плазмиды Escherichia coli . J. Biomed. Sci. 9, 246–252. DOI: 10.1159 / 000059425
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Венсинк, П. К., Финнеган, Д. Дж., Донельсон, Дж. Э. и Хогнесс, Д. С. (1974). Система для картирования последовательностей ДНК в хромосомах Drosophila melanogaster . Cell 3, 315–325. DOI: 10.1016 / 0092-8674 (74) -2
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Уэстон, А., Браун, М. Г., Перкинс, Х. Р., Сондерс, Дж. Р. и Хамфрис, Г. О. (1981). Трансформация Escherichia coli плазмидой дезоксирибонуклеиновой кислоты: индуцированное кальцием связывание дезоксирибонуклеиновой кислоты с целыми клетками и изолированными фракциями мембран. J. Bacteriol. 145, 780–787.
PubMed Аннотация | Google Scholar
Рабочий процесс бактериальной трансформации — 4 основных шага | Thermo Fisher Scientific
Бактериальная трансформация — ключевой этап молекулярного клонирования, целью которого является получение нескольких копий рекомбинантной молекулы ДНК.Предыдущие этапы создания рекомбинантных плазмид описаны в традиционных основах клонирования и включают в себя вставку интересующей последовательности ДНК в основу вектора. При трансформации ДНК (обычно в форме плазмиды) вводится в компетентный штамм бактерий, чтобы бактерии затем могли реплицировать интересующую последовательность в количествах, подходящих для дальнейшего анализа и / или манипуляции.
Четыре ключевых этапа бактериальной трансформации:
Рисунок 1.Ключевые этапы процесса бактериальной трансформации: (1) подготовка компетентных клеток, (2) трансформация клеток, (3) восстановление клеток и (4) посев клеток.
Грамотная подготовка клеток
E. coli — наиболее распространенный вид бактерий, используемый на этапе трансформации рабочего процесса клонирования. Поскольку естественная компетентность E. coli очень низкая или даже отсутствует, клетки необходимо сделать компетентными для трансформации с помощью теплового шока или электропорации.
Протоколы приготовления компетентных клеток различаются в зависимости от того, будет ли достигнута трансформация с помощью теплового шока или электропорации. В любом случае одна свежая колония желаемого штамма берется из чашки с агаром и инокулируется в жидкую среду для заквасочной культуры (, рис. 2, ). Эту заквасочную культуру и последующую более крупную культуру тщательно контролируют на предмет активного роста путем непрерывного измерения оптической плотности при 600 нм (OD 600 ). Для достижения высокой эффективности трансформации очень важно, чтобы рост клеток происходил в фазе среднего логарифма во время сбора урожая, что обычно происходит при OD 600 между 0.4 и 0,9, с оптимальным значением в зависимости от объема культуры, штамма и протокола. На всех этапах необходимо использовать стерильные инструменты и лабораторную посуду, среды и реагенты, где это необходимо или необходимо. После сбора клеток для дальнейшей обработки рекомендуется хранить все образцы, реагенты и оборудование при 0–4 ° C, чтобы повысить жизнеспособность клеток и сохранить эффективность трансформации.
Собранные клетки затем обрабатывают в соответствии с методом трансформации, будь то тепловой шок или электропорация ( Рисунок 2 ).
- Трансформация тепловым шоком: Компетентные клетки получают химическим путем путем инкубации клеток в хлориде кальция (CaCl 2 ), чтобы сделать клеточную мембрану более проницаемой [1,2]. Для дальнейшего повышения компетентности Ca 2+ может быть дополнен или заменен другими катионами и реагентами, такими как марганец (Mn 2+ ), калий (K + ), кобальт ([Co (NH₃) ₆] 3+ ), рубидий (Rb + ), диметилсульфоксид (ДМСО) и / или дитиотреитол (DTT), как описано Hannah et al.(1983) [3].
- Электропорация: Собранные клетки несколько раз промывают ледяной деионизированной водой путем повторного гранулирования и ресуспендирования для удаления солей и других компонентов, которые могут мешать электропорации. После 3-4 промываний клетки окончательно осаждают и ресуспендируют в 10% глицерине для хранения [4].
Рисунок 2. Получение химически компетентных и электрокомпетентных клеток.
После приготовления компетентные клетки должны быть оценены на эффективность трансформации, разделены на аликвоты до небольших объемов, чтобы минимизировать циклы замораживания / оттаивания, и храниться при соответствующей температуре для поддержания жизнеспособности.Эффективность трансформации компетентных клеток обычно измеряется по поглощению субнасыщающих количеств суперспиральной интактной плазмиды (например, 10-500 пг ДНК pUC). Результаты выражаются в количестве образованных колоний (трансформантов) или колониеобразующих единиц (КОЕ) на микрограмм использованной плазмидной ДНК (КОЕ / мкг) (см. Посев клеток).
Для хранения рекомендуется аликвотирование подготовленных клеток в одноразовых объемах в микроцентрифужных пробирках с завинчивающейся крышкой, поскольку каждый цикл замораживания / оттаивания снижает эффективность трансформации примерно наполовину.Компетентные клетки должны оставаться стабильными в течение примерно 6–12 месяцев при хранении при –70 ° C с минимальными колебаниями температуры. Клетки , а не следует замораживать или хранить в жидком азоте, поскольку такая практика резко снижает жизнеспособность.
Для единообразия и экономии времени готовых компетентных клеток доступны в готовых к использованию форматах из коммерческих источников. Эти компетентные клетки проходят контроль качества и тестируются на соответствие спецификациям по эффективности трансформации и генотипам.Эти препараты минимизируют вариабельность от партии к партии и значительно упрощают эффективное размножение клонированной ДНК.
Трансформация
Двумя наиболее популярными методами бактериальной трансформации являются (1) тепловой шок химически подготовленных компетентных клеток (химическая трансформация) и (2) электропорация электрокомпетентных клеток. Выбор зависит от требуемой эффективности преобразования, экспериментальных целей и доступных ресурсов (см. Выбор компетентных ячеек).Когда они готовы к стадии трансформации, компетентные клетки следует разморозить на льду и осторожно обработать, чтобы сохранить жизнеспособность. Клетки можно смешивать осторожным встряхиванием, постукиванием или пипетированием, но следует избегать встряхивания.
Обзор химического преобразования
Изучите основы трансформации, два типа компетентных клеток, способы проведения химической трансформации и советы по устранению неполадок.
Примечание: Отрицательный и положительный контроли должны быть включены в этап преобразования, чтобы оценить успех экспериментальной процедуры.
| Контроль | Компоненты образца | Оценка |
|---|---|---|
| Отрицательный |
| |
| Положительный |
| |
| Экспериментальная |
|
С химической трансформацией , химически компетентных клеток смешивают с плазмидной ДНК и ненадолго подвергают воздействию повышенной температуры, процесс, известный как тепловой шок (, рис. 3A, ).Сначала клетки инкубируют с ДНК на льду в течение 5–30 минут в полипропиленовой пробирке. Следует избегать пробирок из полистирола, поскольку ДНК может прилипать к поверхности, снижая эффективность трансформации. Традиционно для достижения наилучших результатов использовались трубы с круглым дном 17 x 100 мм. Использование микроцентрифужных пробирок объемом 1,5 мл может привести к плохому распределению тепла из-за меньшего отношения поверхности к объему суспензии клеток, что может снизить эффективность трансформации на 60–90%, особенно для клеток с более высокой эффективностью.
Для успешной химической трансформации рекомендуется 50–100 мкл компетентных клеток и 1–10 нг ДНК. Когда смесь для лигирования используется в качестве трансформирующей ДНК (часто достаточно 1–5 мкл), очистка перед химической трансформацией обычно не требуется. Важно отметить, что смеси для лигирования могут приводить к эффективности трансформации всего 1–10% по сравнению с трансформацией с суперспиральной интактной плазмидной ДНК.
Тепловой шок выполняется при 37–42 ° C в течение 25–45 секунд в зависимости от используемого бактериального штамма и ДНК.Для меньших объемов ячеек в меньших трубках интервал теплового шока, который зависит от отношения поверхности к объему суспензии ячеек, должен быть уменьшен. Затем клетки, подвергшиеся тепловому шоку, возвращают в лед на ≥2 минут перед следующим этапом (, рис. 3A, ).
Рис. 3. Бактериальная трансформация с использованием (A) химически компетентных клеток и теплового шока и (B) электрокомпетентных клеток и электропорации.
Электропорация включает использование электропоратора для воздействия на компетентные клетки и ДНК короткого импульса высоковольтного электрического поля (, рис. 3В, ).Считается, что эта обработка вызывает временные поры в клеточных мембранах, которые позволяют ДНК проникать в клетки (, рис. 4, ). Наиболее распространенным типом электрического импульса при бактериальной трансформации является экспоненциальный спад, при котором заданное напряжение прикладывается и затухает в течение нескольких миллисекунд, что называется постоянной времени (, рис. 4A, ). Приложенное напряжение определяется напряженностью поля (В / см), где V — начальное пиковое напряжение, а см — измерение зазора между электродами используемой кюветы.Обычно для электропорации бактерий используются кюветы диаметром 0,1 см (объемом 20–80 мкл) и требуется напряженность поля> 15 кВ / см.
Рис. 4. (A) Экспоненциальное затухание электрического импульса. (B) Процесс электропорации.
Одной из основных проблем электропорации является искрение или электрический разряд, который может снизить жизнеспособность клеток и эффективность трансформации. Возникновение дуги часто возникает в результате электропорации в проводящих буферах, например, содержащих MgCl 2 и фосфаты.
Стратегии предотвращения образования дуги включают следующее:
- Минимизируйте ионную силу растворов ДНК и буферов электропорации. Смеси ДНК для лигирования должны быть очищены на колонке и ресуспендированы в воде или буфере ТЕ для удаления белков и солей перед электропорацией.
- Сохраняйте объем раствора ДНК на уровне не более 5% от общего объема суспензии клеток (например, 2 мкл ДНК на 40 мкл клеток).
- Избегайте уноса агара во время подготовки электрокомпетентных клеток.
- Убедитесь, что в кювете для электропорации нет пузырьков воздуха.
- Выдавите клетки прямо на дно кюветы.
После трансформации неиспользованные компетентные клетки (полученные для любого метода) можно повторно заморозить. Однако это снизит эффективность трансформации примерно на 50% для каждого цикла замораживания / оттаивания. Для достижения наилучших результатов разделите клетки после начальной подготовки на одноразовые объемы, чтобы свести к минимуму замораживание и оттаивание. Одноразовый формат коммерчески доступен для трансформации и восстановления в одной и той же пробирке и для исключения необходимости замораживания и оттаивания клеток.Чтобы повторно заморозить неиспользованные клетки, быстро заморозьте их на бане сухой лед / этанол в течение 5 минут и храните при –70 ° C. Избегайте замораживания или хранения клеток в жидком азоте, так как это резко снижает жизнеспособность.
Период восстановления клеток
После теплового шока или электропорации трансформированные клетки культивируют в жидкой среде без антибиотиков в течение короткого периода, чтобы дать возможность начать экспрессию гена (ов) устойчивости к антибиотикам из приобретенной плазмиды (, фиг. 5, ).Этот шаг улучшает жизнеспособность клеток и эффективность клонирования. Для клеток с электропорацией рекомендуется как можно скорее выращивать клетки, поскольку буферы для электропорации не предназначены для длительного выживания клеток.
На стадии восстановления трансформированные клетки культивируют в 1 мл предварительно нагретого S.O.C. среда при 37 ° C со встряхиванием при 225 об / мин в течение 1 часа. S.O.C. среда, содержащая глюкозу и MgCl 2 , рекомендуется для максимальной эффективности трансформации [3]. Использование S.O.C. среда вместо бульона Lennox L (бульон LB) может увеличивать образование трансформированных колоний в 2-3 раза [5].Штаммы для размножения векторов бактериофага M13 не требуют этого этапа.
Рисунок 5. Рост клеток на этапе восстановления.
Советы по нанесению покрытия на ячейки
После выращивания в S.O.C. среды, клетки высевают на агар LB с подходящим антибиотиком (ами) или другими агентами для идентификации и выделения успешных трансформантов. Например, если необходимо выполнить скрининг на синий / белый, в чашку с агаром должны быть включены X-Gal и IPTG. Избегайте использования чашек с агаром старше нескольких недель (или дней в некоторых случаях), чтобы убедиться, что антибиотик активен.Перед посевом клеток чашки следует предварительно нагреть до благоприятной температуры роста и избежать конденсации, чтобы предотвратить контаминацию и смешивание колоний.
Посеянное количество клеток должно дать достаточное (а также не слишком большое) количество отдельных, отличных колоний для дальнейшего скрининга. Клетки, культивируемые в S.O.C. Среду можно осаждать центрифугированием в течение 5 минут при 600–800 x г и ресуспендировать в меньшем объеме для посева. Для посева на 100-миллиметровую пластину обычно хорошо подходит 100–200 мкл клеточной суспензии.Если ожидается очень мало колоний, можно высеять всю клеточную суспензию. Однако, если ожидается очень большое количество колоний, суспензию клеток можно разбавить до 1: 100 в S.O.C. среды перед посевом, чтобы избежать образования бактериального газона.
Рис. 6. Пипетку Пастера можно превратить в одноразовую самодельную «хоккейную клюшку» или L-образный распределитель.
Равномерное распределение клеток на чашке с агаром имеет решающее значение для анализа колоний.Стерильная хоккейная клюшка или L-образный распределитель клеток обычно используется для распределения клеточной суспензии при осторожном вращении планшета (, рис. 6, 7A, ). Избегайте прокалывания поверхности агара при распределении клеток. В качестве альтернативы можно использовать автоклавированные стеклянные шарики (диаметром 4 мм) для распределения клеток. В этом подходе от 10 до 20 шариков помещают на планшет после нанесения суспензии клеток, и планшет осторожно вращают так, чтобы суспензия клеток распределялась по шарикам ( Рисунок 7B ).Клетки необходимо быстро разложить до высыхания жидкой суспензии. После распределения дайте планшету высохнуть перед инкубацией в течение ночи при 37 ° C в перевернутом положении.
Рис. 7. Два распространенных метода нанесения покрытия. ( A ) Распространение стерильной хоккейной клюшкой. ( B ) Распространение стерильными стеклянными шариками 4 мм и осторожным вращением чашки.
На следующий день чашки с культурой исследуют на образование колоний. Следует избегать продолжительной инкубации, поскольку она часто приводит к слиянию больших колоний и появлению более мелких, чувствительных к антибиотикам окружающих колоний (так называемых сателлитных колоний ) из-за разложения антибиотиков вокруг больших колоний.
Чтобы рассчитать эффективность трансформации, разделите количество трансформантов на количество добавленной ДНК и фактор разведения клеток (если выполнено), используя следующую формулу:
С лигированной ДНК количество ДНК, добавленной к клеткам, может также может быть определено из настройки реакции лигирования, разведения ДНК (если выполнено) и объема ДНК для трансформации, используя следующую формулу:
Пример расчета эффективности трансформации
50 нг ДНК лигируют в 20 мкл реакции.После лигирования реакционную смесь разбавляют в 2 раза и 5 мкл разбавленной смеси для лигирования добавляют к 100 мкл компетентных клеток для трансформации.
ДНК, добавленная к клеткам = (0,05 мкг / 20 мкл) x 1/2 x 5 мкл = 0,00625 мкг
После трансформации суспензию клеток разбавляют в 5 раз и высевают на чашки по 200 мкл разведенных клеток. После инкубации в течение ночи образуется 300 колоний.
Эффективность трансформации = (300 КОЕ / 0,00625 мкг) x (100 мкл / 200 мкл) x 5 = 1.2 x 10 5 КОЕ / мкг
Колонии необходимо подвергнуть дополнительному скринингу на наличие желаемой плазмиды и при необходимости исправить последовательность (см. Методы скрининга колоний). После подтверждения желаемые колонии можно использовать в последующих приложениях, таких как выделение плазмид, субклонирование, трансфекция и экспрессия белка.
Рекомендации
Естественная генетическая трансформация: распространенность, механизмы и функции
Сравнительный анализ геномов прокариот показывает, что приобретение генетического материала посредством латерального переноса генов было основной движущей силой в эволюции этих организмов.Однако обмен генетическим материалом может ускорить эволюцию только в том случае, если доноры и получатели будут использовать одну и ту же систему для кодирования, хранения и обработки генетической информации. Следовательно, прокариотический «пол» должен был сыграть значительную роль в сохранении почти универсальности генетического кода. При исследовании 88 геномов прокариот, процент латерально переносимых генов колеблется от 0 до 22% у бактерий и 5-15% у видов архей, соответственно [50]. Эти результаты, которые, вероятно, представляют собой заниженную оценку, показывают, что латеральный перенос ДНК вместе с мутациями, потерей генов и дублированием существующих генов формирует геномы организмов в обеих сферах жизни.Согласно литературным данным, бактерии и археи используют одни и те же механизмы, то есть конъюгацию, трансдукцию и естественную трансформацию, для приобретения экзогенной ДНК [86], [87]. Это, несомненно, верно для конъюгации и трансдукции, но вносит ли естественная трансформация вообще вклад в латеральный перенос генов у архей, в настоящее время неизвестно из-за отсутствия данных. Напротив, естественная трансформация, которая определяется как активное поглощение и наследуемая интеграция внеклеточной ДНК, была тщательно изучена на отдельных видах бактерий, таких как Streptococcus pneumoniae , Neisseria spp., Bacillus subtilis и Haemophilus influenzae [24], [30], [60], [61], [120]. В отличие от конъюгации и трансдукции, перенос ДНК путем естественной трансформации инициируется клеткой-реципиентом. Таким образом, в то время как конъюгация и трансдукция зависят от внехромосомных генетических элементов, способствующих их собственному поддержанию и распространению, естественная трансформация является частью нормальной физиологии компетентной бактерии и, следовательно, может считаться уникально адаптированной к потребностям хозяина.
В этом обзоре мы даем краткий обзор филогенетического распределения естественно трансформируемых бактерий, обобщаем текущие знания о механизмах захвата и высвобождения ДНК и обсуждаем, как естественная трансформация может принести пользу бактериям, обладающим этим свойством.
Влияние этапа теплового шока на эффективность бактериальной трансформации
Mol Biol Res Commun. 2016 Dec; 5 (4): 257–261.
Марал Рахимзаде
1 Кафедра нанобиотехнологии, Школа биологических наук, Университет Тарбиат Модарес, Тегеран, Иран
Маджид Садегизаде
2 Кафедра молекулярной генетики, Тарбихатский университет биологических наук Иран
Farhood Najafi
3 Департамент смол и добавок, Институт науки и технологии цвета, Тегеран, Иран
Сейед Араб
4 Департамент биофизики, Школа биологических наук, Университет Тарбиат Модарес, Тегеран, Иран
Хамид Мобашери
5 5) Лаборатория биофизики мембран, Институт биохимии и биофизики, Тегеранский университет, Тегеран, Иран
1 Кафедра нанобиотехнологии, Школа биологических наук, Тегеранский университет Тарбиат Модарес Иран
2 Департамент молекулярной генетики, Школа биологических наук, Университет Тарбиат Модарес, Тегеран, Иран
3 Отделение смол и добавок, Институт науки и технологий цвета, Тегеран, Иран
4 Отделение биофизики, Школа биологических наук, Университет Тарбиат Модарес , Тегеран, Иран
5 5) Лаборатория биофизики мембран, Институт биохимии и биофизики, Тегеранский университет, Тегеран, Иран
* Автор для корреспонденции: Департамент молекулярной генетики, Школа биологических наук, Университет Тарбиат Модарес , Тегеран, Иран, тел .: +98 21-82884409, факс: +98 21-82884717, E.mail: [email protected]
Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License (http://creativecommons.org/licenses/by/3.0/), которая разрешает неограниченное использование, распространение, и воспроизведение на любом носителе при условии правильного цитирования оригинальной работы. Эта статья цитировалась в других статьях PMC.Abstract
CaCl 2 Обработка с последующим тепловым шоком является наиболее распространенным методом искусственного преобразования.Здесь клетки трансформировали с использованием обработки CaCl 2 либо с тепловым шоком (стандартный протокол), либо без теплового шока (лабораторный протокол), чтобы понять разницу в эффективности трансформации. Штамм BL21 Escherichia coli ( E. coli ) был чувствительным при обработке CaCl 2 . Некоторые клетки хранили при -80 o ° C, а другие — при 4 ° C. После этого чувствительные клетки трансформировали с использованием стандартного или лабораторного протокола.Эффективность трансформации между ячейками, испытавшими тепловой шок, и те, на которые не повлиял тепловой шок, была почти одинаковой. Более того, независимо от протокола трансформации, клетки, хранящиеся при 4 ˚C, трансформировались более эффективно по сравнению с клетками, которые хранились при -80 o C.
Ключевые слова: E. coli , Искусственная трансформация, тепловой шок , Эффективность трансформации
ВВЕДЕНИЕ
Бактерии могут естественным образом получать новую общую информацию с помощью 3 различных механизмов; конъюгация, трансдукция и трансформация [1].Перенос ДНК напрямую от одного организма к другому называется конъюгацией, но при трансдукции перенос ДНК происходит с помощью бактериофага. Во время трансформации голая ДНК связывается с поверхностью клетки и проходит через комплекс стенка-мембрана [2]. Трансформация бактерий может происходить естественным или искусственным путем. Естественная трансформация — это редкий механизм, используемый некоторыми бактериальными клетками для поглощения ДНК из окружающей среды. При искусственной трансформации бактериальные клетки должны быть чувствительны в определенных лабораторных условиях до трансформации.Есть два основных метода искусственной трансформации бактерий; Обработка CaCl 2 с последующим коротким тепловым шоком и электропорацией [3, 4]. Любой из методов был изменен за последнее столетие для повышения эффективности трансформации.
Точный механизм того, как обработка CaCl 2 может облегчить перенос ДНК из внеклеточной среды в цитоплазму, все еще не раскрыт. Предполагается, что молекулы ДНК могут абсорбироваться на поверхности клетки с помощью двухвалентного катиона Ca 2+ и ступень теплового шока делает возможным попадание ДНК в цитозоль [2].
В этом исследовании бактерий E. coli трансформировали двумя способами; (1) обработка CaCl 2 с последующей стадией теплового шока и (2) обработка CaCl 2 без использования стадии теплового шока. Эффективность трансформации рассчитывалась для обоих методов. Кажется, что этап теплового шока не может иметь решающего значения для протокола трансформации.
МАТЕРИАЛЫ И МЕТОДЫ
Для трансформации использовали штамм BL21 бактерии E. coli .Вектор pEGFP-N1, первоначально приобретенный в Clontech Laboratories, Inc. (США). Вектор устойчив к канамицину и содержит ген усиленного зеленого флуоресцентного белка (EGFP).
E. coli бактерий выращивали в среде Luria Bertani (LB). 15 мкл культивированного в течение ночи E. coli инокулировали в 25 мл бульона LB и инкубировали на шейкере при 37 ° C и 250 об / мин до тех пор, пока оптическая плотность суспензии при 600 нм не достигала диапазона 0,4-0,7. Затем суспензии выдерживали на льду 30 мин [5].После этого этапа бактерии осаждали с помощью центрифуги Dragon Lab (D3024R, Dragon Laboratory Instruments Limited, Китай) при 1717 × г в течение 20 минут при 4 ° C. Бактериальный осадок ресуспендировали в 12,5 мл CaCl 2 (100 мМ) и помещали на лед на 30 мин. Затем клеточные суспензии центрифугировали, как указано выше, и осадки растворяли в 25 мл CaCl 2 (100 мМ). На этом этапе суспензии держат на льду всю ночь. Снова бактерии осаждали при 1717 × г в течение 20 минут при 4 ˚C, и осадки повторно суспендировали в 2250 мкл CaCl 2 (100 мМ) и 750 мкл глицерина 60% и хранили при -80 ˚C и / или 4 ˚C для дальнейших экспериментов.
Преобразования выполняли с использованием стандартного или лабораторного протокола следующим образом; 100 мкл размороженных компетентных бактерий и 0,65 нг ДНК pEGFP-N1 (выделенных из штамма DH5-alpha E. coli с помощью набора для мини-экстракции плазмид AccuPrep® от Bioneer и проверенных с помощью гель-агарозного электрофореза) добавляли в предварительно охлажденную пробирку и аккуратно пипетировать. По стандартному протоколу суспензии выдерживали на льду в течение 30 минут, после чего смесь нагревали при 42 ° C в течение 45 секунд.После этапа теплового шока пробирки выдерживали на льду еще 5 мин. Затем добавляли 1 мл бульона LB и готовили серийно разведенные суспензии, которые инкубировали в шейкере при 37 ° C в течение 2 часов. Но в нашем лабораторном протоколе этапы льда и теплового шока были полностью опущены, и 1 мл бульона LB был немедленно добавлен к смеси ДНК и бактериальных клеток (при 25 ˚C), а суспензия была серийно разбавлена и инкубирована в шейкере при 250 ° C. об / мин в течение 2 часов при 37 ˚C. В конце обоих протоколов клетки центрифугировали при 1717 × g в течение 5 мин.и осадки ресуспендировали в 100 мкл LB и наносили штрихами на устойчивый к канамицину планшет.
Эксперименты проводились четырехкратно. Данные представлены как среднее ± стандартное отклонение. Непарный t-тест был проведен для того, чтобы понять значимость различий. Двусторонним значениям P было присвоено менее 0,05. Для анализа использовалось программное обеспечение GraphPad Prism 5.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Трансформацию проводили с использованием клеток, хранимых при 4 ˚C (свежие) или -80 ˚C как по стандартному, так и по лабораторному протоколу.Клетки были успешно трансформированы стандартным и лабораторным протоколом. Количество бактерий, способных поглощать молекулы ДНК, определяется как эффективность трансформации (TE) [2]. TE рассчитывалось по уравнению 1.
ТЕ = количество колониеобразующих единиц (КОЕ) количество (в мкг) плазмидной ДНК
В целом ТЕ свежих клеток (выдержанных при 4 ˚C) было больше, чем те, которые хранились в течение более длительного периода при -80 o C. Примечательно, что разница между лабораторным и стандартным протоколом была довольно низкой, и бактерии можно было трансформировать без применения теплового шока.Эффективность трансформации отображается в формате. Таблички успешной трансформации представлены в.
Эффективность трансформации в различных условиях
Планшеты для успешной бактериальной трансформации с использованием стандартного и лабораторного протокола (образцы были разбавлены в 100 раз
Было высказано предположение, что стадия теплового шока может облегчить проникновение в ДНК, но все еще недостаточно подсказок. Panja et al. (2006) сообщили, что ступенька теплового импульса вызывает снижение мембранного потенциала [2].Внутренний потенциал клетки становится менее отрицательным в результате уменьшения мембранного потенциала, поэтому отрицательная ДНК может легче проникать в цитозоль [2].
Хотя точная роль ионов Ca 2+ еще не известна, считается, что Ca 2+ может развивать взаимодействие между молекулами ДНК и LPS (липополисахаридом) внешней мембраны. Кроме того, ионы Ca 2+ могут изменять физико-химические свойства липидов и вызывать фазовый переход фосфатидилглицерина и ЛПС [6-8].Кроме того, двухвалентные катионы Ca 2+ усиливают структурные изменения в бислоев фосфатидилхолин-кардиолипин, что приводит к увеличению проницаемости [6, 9-11].
Наши результаты показывают, что ионы Ca 2+ играют более важную роль в искусственной трансформации. Кроме того, кажется, что способность Ca 2+ увеличивать проницаемость мембраны больше, чем тепловой шок. В то же время следует отметить, что в протоколе лаборатории клетки подверглись кратковременному тепловому шоку (0 → 25 → 37 ˚C), которого оказалось достаточно для переноса ДНК.Чтобы понять точный механизм превращения ионов Ca 2+ и теплового шока в стадию превращения, необходимо провести дополнительные исследования.
Выражение признательности
Этот исследовательский проект финансировался за счет гранта факультета биологических наук Университета Тарбиат Модарес, Тегеран, Иран. Авторы заявляют об отсутствии конфликта интересов.
Конфликт интересов:
Авторы заявляют об отсутствии конфликта интересов.